Տարբեր տեսակի հյուսվածքների հյուսվածաբանական կառուցվածքը. Հյուսվածքաբանության հիմունքներ. Հյուսվածքների դասակարգում. էպիթելային հյուսվածք. Շարակցական հյուսվածքի. Բջջ - կյանքի տարրական միավոր
ՀԱՏՄԱԲԱՆՈՒԹՅՈՒՆ
գիտություն, որն ուսումնասիրում է կենդանիների հյուսվածքները։ Հյուսվածքը բջիջների խումբ է, որոնք նման են ձևով, չափսով և ֆունկցիայով և իրենց նյութափոխանակության արտադրանքներով: Բոլոր բույսերի և կենդանիների, բացառությամբ առավել պարզունակների, մարմինը բաղկացած է հյուսվածքներից, իսկ բարձրակարգ բույսերում և բարձր կազմակերպված կենդանիների մոտ հյուսվածքներն առանձնանում են կառուցվածքի մեծ բազմազանությամբ և դրանց արտադրանքի բարդությամբ. զուգակցվելով միմյանց հետ՝ տարբեր հյուսվածքներ կազմում են մարմնի առանձին օրգաններ։ Հյուսվածքաբանությունը կենդանիների հյուսվածքների ուսումնասիրությունն է. Բուսական հյուսվածքների ուսումնասիրությունը սովորաբար կոչվում է բույսերի անատոմիա: Հյուսվածքաբանությունը երբեմն կոչվում է մանրադիտակային անատոմիա, քանի որ այն ուսումնասիրում է մարմնի կառուցվածքը (ձևաբանությունը) միկրոսկոպիկ մակարդակով (հյուսվածքի շատ բարակ հատվածները և առանձին բջիջները ծառայում են որպես հյուսվածքաբանական հետազոտության առարկա): Չնայած այս գիտությունը հիմնականում նկարագրական է, նրա խնդիրը ներառում է նաև այն փոփոխությունների մեկնաբանումը, որոնք տեղի են ունենում հյուսվածքներում նորմալ և պաթոլոգիական պայմաններում: Հետևաբար, հյուսվածաբանը պետք է լավ իմանա, թե ինչպես են հյուսվածքները ձևավորվում սաղմնային զարգացման գործընթացում, ինչպիսի՞ն է դրանց աճի կարողությունը հետսաղմնային շրջանում և ինչպես են դրանք ենթարկվում փոփոխությունների տարբեր բնական և փորձարարական պայմաններում, այդ թվում՝ ծերացման և ծերացման ընթացքում։ դրանց բաղկացուցիչ բջիջների մահը. Հյուսվածքաբանության պատմությունը՝ որպես կենսաբանության առանձին ճյուղ, սերտորեն կապված է մանրադիտակի ստեղծման և դրա կատարելագործման հետ։ M. Malpighi (1628-1694) կոչվում է «միկրոսկոպիկ անատոմիայի հայր», հետևաբար՝ հյուսվածաբանություն: Հյուսվածքաբանությունը հարստացել է բազմաթիվ գիտնականների կողմից իրականացված կամ ստեղծած հետազոտությունների դիտարկումներով և մեթոդներով, որոնց հիմնական հետաքրքրությունները կենդանաբանության կամ բժշկության բնագավառում են: Դրա մասին է վկայում հյուսվածաբանական տերմինաբանությունը, որը հավերժացրել է նրանց անունները իրենց առաջին նկարագրած կառույցների կամ ստեղծած մեթոդների անուններով. Ներկայումս լայն տարածում են գտել պատրաստուկների պատրաստման մեթոդները և դրանց մանրադիտակային հետազոտությունը, ինչը հնարավորություն է տալիս ուսումնասիրել առանձին բջիջներ։ Այս մեթոդները ներառում են սառեցված հատվածի տեխնիկան, ֆազային կոնտրաստային մանրադիտակը, հիստոքիմիական անալիզը, հյուսվածքների կուլտուրան, էլեկտրոնային մանրադիտակը; վերջինս թույլ է տալիս մանրամասն ուսումնասիրել բջջային կառուցվածքները (բջջաթաղանթներ, միտոքոնդրիաներ և այլն)։ Սկանավորող էլեկտրոնային մանրադիտակի միջոցով հնարավոր եղավ բացահայտել բջիջների և հյուսվածքների ազատ մակերեսների հետաքրքիր եռաչափ կոնֆիգուրացիան, որը հնարավոր չէ տեսնել սովորական մանրադիտակի տակ:
Հյուսվածքների ծագումը.Բեղմնավորված ձվաբջջից սաղմի զարգացումը տեղի է ունենում բարձրակարգ կենդանիների մոտ բջիջների բազմակի բաժանման (ջախջախման) արդյունքում. այս դեպքում գոյացած բջիջները աստիճանաբար բաշխվում են իրենց տեղերում՝ ապագա սաղմի տարբեր հատվածներում։ Սկզբում սաղմնային բջիջները նման են միմյանց, սակայն դրանց քանակի աճի հետ նրանք սկսում են փոխվել՝ ձեռք բերելով բնորոշ հատկանիշներ և որոշակի հատուկ գործառույթներ կատարելու ունակություն։ Այս գործընթացը, որը կոչվում է տարբերակում, ի վերջո հանգեցնում է տարբեր հյուսվածքների ձեւավորմանը: Ցանկացած կենդանու բոլոր հյուսվածքները ծագում են երեք սկզբնական սաղմնային շերտերից. 1) արտաքին շերտը կամ էկտոդերմը. 2) ամենաներքին շերտը կամ էնդոդերմը. և 3) միջին շերտը կամ մեզոդերմը: Այսպիսով, օրինակ, մկանները և արյունը մեզոդերմի ածանցյալներն են, աղիքային տրակտի լորձաթաղանթը զարգանում է էնդոդերմայից, իսկ էկտոդերմը ձևավորում է ներքին հյուսվածքներ և նյարդային համակարգը:
Տես նաև ՍԱՂՄԲԱՆՈՒԹՅՈՒՆ։
գործվածքների հիմնական տեսակները.Հյուսվածքաբանները սովորաբար առանձնացնում են մարդկանց և բարձր կենդանիների չորս հիմնական հյուսվածքները՝ էպիթելային, մկանային, միացնող (ներառյալ արյունը) և նյարդային: Որոշ հյուսվածքներում բջիջներն ունեն մոտավորապես նույն ձևն ու չափը և այնքան ամուր կից են միմյանց, որ դրանց միջև միջբջջային տարածություն չկա կամ գրեթե բացակայում է. նման հյուսվածքները ծածկում են մարմնի արտաքին մակերեսը և երեսպատում նրա ներքին խոռոչները։ Մյուս հյուսվածքներում (ոսկոր, աճառ) բջիջներն այնքան էլ խիտ չեն լցված և շրջապատված են իրենց արտադրած միջբջջային նյութով (մատրիքսով)։ Նյարդային հյուսվածքի (նեյրոնների) բջիջներից, որոնք կազմում են ուղեղը և ողնուղեղը, հեռանում են երկար գործընթացներ, որոնք ավարտվում են բջջային մարմնից շատ հեռու, օրինակ՝ մկանային բջիջների հետ շփման կետերում։ Այսպիսով, յուրաքանչյուր հյուսվածք կարող է տարբերվել մյուսներից բջիջների գտնվելու վայրի բնույթով: Որոշ հյուսվածքներ ունեն սինցիցիալ կառուցվածք, որի դեպքում մեկ բջջի ցիտոպլազմիկ պրոցեսները անցնում են հարևան բջիջների նմանատիպ գործընթացների մեջ. նման կառուցվածք նկատվում է բողբոջային մեզենխիմում, թուլացած շարակցական հյուսվածքում, ցանցանման հյուսվածքում և կարող է առաջանալ նաև որոշ հիվանդությունների ժամանակ։ Շատ օրգաններ կազմված են մի քանի տեսակի հյուսվածքներից, որոնք կարելի է ճանաչել իրենց բնորոշ մանրադիտակային կառուցվածքով։ Ստորև բերված է բոլոր ողնաշարավորների մոտ հայտնաբերված հյուսվածքների հիմնական տեսակների նկարագրությունը: Անողնաշարավորները, բացառությամբ սպունգների և կոելենտերատների, ունեն նաև մասնագիտացված հյուսվածքներ, որոնք նման են ողնաշարավորների էպիթելային, մկանային, միացնող և նյարդային հյուսվածքներին։
էպիթելային հյուսվածք.Էպիթելը կարող է բաղկացած լինել շատ հարթ (թևավոր), խորանարդաձև կամ գլանաձև բջիջներից։ Երբեմն այն բազմաշերտ է, այսինքն. բաղկացած բջիջների մի քանի շերտերից; նման էպիթելը ձևավորում է, օրինակ, մարդու մաշկի արտաքին շերտը։ Մարմնի այլ մասերում, օրինակ՝ ստամոքս-աղիքային տրակտում, էպիթելը միաշերտ է, այսինքն. նրա բոլոր բջիջները կապված են հիմքում ընկած նկուղային թաղանթին: Որոշ դեպքերում, միաշերտ էպիթելը կարող է թվալ բազմաշերտ. եթե նրա բջիջների երկար առանցքները միմյանց զուգահեռ չեն, ապա թվում է, որ բջիջները գտնվում են տարբեր մակարդակների վրա, թեև իրականում դրանք ընկած են նույնի վրա: նկուղային թաղանթ. Նման էպիթելը կոչվում է բազմաշերտ: Էպիթելի բջիջների ազատ եզրը ծածկված է թարթիչներով, այսինքն. պրոտոպլազմայի բարակ մազերի նման աճը (այդպիսի թարթիչավոր էպիթելի գծեր, օրինակ՝ շնչափող) կամ ավարտվում է «խոզանակի եզրագծով» (բարակ աղիքները ծածկող էպիթելի); այս եզրագիծը բաղկացած է բջջի մակերեսի ուլտրամիկրոսկոպիկ մատների նման ելքերից (այսպես կոչված՝ միկրովիլիներ): Պաշտպանիչ գործառույթներից բացի, էպիթելը ծառայում է որպես կենդանի թաղանթ, որի միջոցով գազերը և լուծված նյութերը ներծծվում են բջիջների կողմից և արտանետվում դեպի արտաքին: Բացի այդ, էպիթելը ձևավորում է մասնագիտացված կառույցներ, ինչպիսիք են գեղձերը, որոնք արտադրում են մարմնի համար անհրաժեշտ նյութեր: Երբեմն գաղտնի բջիջները ցրված են էպիթելային այլ բջիջների մեջ. Օրինակ՝ լորձ արտադրող գավաթային բջիջները ձկների մաշկի մակերեսային շերտում կամ կաթնասունների աղիների լորձաթաղանթում:

Մկանային.Մկանային հյուսվածքը մնացածից տարբերվում է կծկվելու ունակությամբ: Այս հատկությունը պայմանավորված է մկանային բջիջների ներքին կազմակերպմամբ, որոնք պարունակում են մեծ թվով ենթամանրադիտակային կծկվող կառույցներ: Գոյություն ունեն երեք տեսակի մկաններ՝ կմախքային, որը նաև կոչվում է գծավոր կամ կամավոր; հարթ կամ ակամա; սրտի մկանները, որոնք գծավոր, բայց ակամա. Հարթ մկանային հյուսվածքը բաղկացած է spindle ձեւավորված mononuclear բջիջներից: Զոլավոր մկանները ձևավորվում են բազմամիջուկային երկարաձգված կծկվող միավորներից՝ բնորոշ լայնակի շերտավորմամբ, այսինքն. երկար առանցքին ուղղահայաց թեթև և մուգ շերտեր: Սրտի մկանը բաղկացած է միամիջուկային բջիջներից, որոնք կապված են ծայրից ծայր և ունի լայնակի գծավոր; մինչդեռ հարևան բջիջների կծկվող կառուցվածքները միացված են բազմաթիվ անաստոմոզներով՝ կազմելով շարունակական ցանց։

Շարակցական հյուսվածքի.Գոյություն ունեն կապի հյուսվածքի տարբեր տեսակներ: Ողնաշարավորների ամենակարևոր աջակից կառույցները բաղկացած են երկու տեսակի շարակցական հյուսվածքից՝ ոսկորից և աճառից։ Աճառային բջիջները (քոնդրոցիտներ) իրենց շուրջն արտազատում են խիտ առաձգական հիմք նյութ (մատրիքս): Ոսկրային բջիջները (օստեոկլաստները) շրջապատված են աղացած նյութով, որը պարունակում է աղի նստվածքներ, հիմնականում՝ կալցիումի ֆոսֆատ։ Այս հյուսվածքներից յուրաքանչյուրի հետևողականությունը սովորաբար որոշվում է հիմնական նյութի բնույթով: Քանի որ մարմինը ծերանում է, ոսկորների հողային նյութում հանքանյութերի պարունակությունը մեծանում է, և այն դառնում է ավելի փխրուն: Փոքր երեխաների մոտ ոսկրի հիմնական նյութը, ինչպես նաև աճառը, հարուստ է օրգանական նյութերով. դրա պատճառով նրանք սովորաբար ունենում են ոչ թե իրական ոսկորների կոտրվածքներ, այլ այսպես կոչված. կոտրվածքներ («կանաչ ճյուղ» տիպի կոտրվածքներ): Ջլերը կազմված են թելքավոր շարակցական հյուսվածքից; նրա մանրաթելերը ձևավորվում են կոլագենից՝ սպիտակուց, որը արտազատվում է ֆիբրոցիտների (ջիլային բջիջների) կողմից։ ճարպային հյուսվածքը գտնվում է մարմնի տարբեր մասերում; Սա շարակցական հյուսվածքի յուրօրինակ տեսակ է՝ բաղկացած բջիջներից, որոնց կենտրոնում կա ճարպի մեծ գնդիկ։

Արյուն.Արյունը շարակցական հյուսվածքի շատ հատուկ տեսակ է. որոշ հյուսվածաբաններ նույնիսկ այն առանձնացնում են որպես անկախ տեսակ: Ողնաշարավորների արյունը բաղկացած է հեղուկ պլազմայից և ձևավորված տարրերից՝ արյան կարմիր բջիջներից կամ հեմոգլոբին պարունակող էրիթրոցիտներից; մի շարք սպիտակ բջիջներ կամ լեյկոցիտներ (նեյտրոֆիլներ, էոզինոֆիլներ, բազոֆիլներ, լիմֆոցիտներ և մոնոցիտներ) և թրոմբոցիտներ կամ թրոմբոցիտներ։ Կաթնասունների մոտ արյան մեջ մտնող հասուն էրիթրոցիտները միջուկներ չեն պարունակում. մնացած բոլոր ողնաշարավորների մեջ (ձկներ, երկկենցաղներ, սողուններ և թռչուններ) հասուն, գործող էրիթրոցիտները պարունակում են միջուկ: Լեյկոցիտները բաժանվում են երկու խմբի՝ հատիկավոր (գրանուլոցիտներ) և ոչ հատիկավոր (ագրանուլոցիտներ)՝ կախված իրենց ցիտոպլազմայում հատիկների առկայությունից կամ բացակայությունից; Բացի այդ, դրանք հեշտ է տարբերել ներկերի հատուկ խառնուրդով ներկելու միջոցով. էոզինոֆիլ հատիկներն այս ներկումով ձեռք են բերում վառ վարդագույն գույն, մոնոցիտների և լիմֆոցիտների ցիտոպլազմը՝ կապտավուն երանգ, բազոֆիլային հատիկներ՝ մանուշակագույն, նեյտրոֆիլային հատիկներ՝ թույլ մանուշակագույն երանգ: Արյան մեջ բջիջները շրջապատված են թափանցիկ հեղուկով (պլազմա), որի մեջ լուծվում են տարբեր նյութեր։ Արյունը թթվածին է մատակարարում հյուսվածքներին, դրանցից հեռացնում է ածխաթթու գազը և նյութափոխանակության արտադրանքը և մարմնի մի մասից մյուսը տեղափոխում սննդանյութեր և արտազատվող արտադրանքներ, ինչպիսիք են հորմոնները: Տես նաև ԱՐՅՈՒՆ։

նյարդային հյուսվածք.Նյարդային հյուսվածքը բաղկացած է բարձր մասնագիտացված բջիջներից՝ նեյրոններից, որոնք կենտրոնացած են հիմնականում ուղեղի և ողնուղեղի գորշ նյութում։ Նեյրոնի (աքսոնի) երկար պրոցեսը ձգվում է միջուկ պարունակող նյարդային բջջի մարմինը երկար հեռավորությունների վրա։ Շատ նեյրոնների աքսոնները կազմում են կապոցներ, որոնք մենք անվանում ենք նյարդեր։ Դենդրիտները հեռանում են նաև նեյրոններից՝ ավելի կարճ պրոցեսներ, սովորաբար բազմաթիվ և ճյուղավորված: Շատ աքսոններ ծածկված են հատուկ միելինային պատյանով, որը կազմված է Շվանի բջիջներից, որոնք պարունակում են ճարպի նմանվող նյութ։ Հարևան Schwann բջիջները բաժանված են փոքր բացերով, որոնք կոչվում են Ranvier հանգույցներ; նրանք ձևավորում են բնորոշ իջվածքներ աքսոնի վրա։ Նյարդային հյուսվածքը շրջապատված է հատուկ տեսակի օժանդակ հյուսվածքով, որը հայտնի է որպես նեյրոգլիա:

Լուգանսկի ազգային ագրարային համալսարան
Բջջաբանություն, սաղմնաբանություն, ընդհանուր հյուսվածաբանություն
(դասախոսական դասընթաց)
Լուգանսկ - 2005 թ
Բջջաբանություն, սաղմնաբանություն, ընդհանուր հյուսվածաբանություն
Դասախոսությունների դասընթացը կազմել է Կենդանիների կենսաբանության ամբիոնի վարիչ, կենսաբանական գիտությունների դոկտոր, պրոֆեսոր Գ.Դ. Կացին։
2-րդ հրատարակություն՝ վերանայված և ընդլայնված։
Դասախոսություններ են պատրաստվում Լուգանսկի ազգային ագրարային համալսարանի զոոբիոտեխնոլոգիական և անասնաբուժական ֆակուլտետի ուսանողների համար: Անկեղծ շնորհակալություն եմ հայտնում կենդանիների կենսաբանության ամբիոնի ասպիրանտ Կրիցյա Յա.Պ. և լաբորատորիայի վարիչ Եսաուլենկո Վ.Պ. նյութը հրապարակման պատրաստելու հարցում նրանց օգնության համար։
Ներածություն հիստոլոգիայի
1. Հյուսվածքաբանության առարկան և նրա տեղը կենսաբանական և անասնաբուժական գիտությունների համակարգում:
2. Մանրադիտակային հետազոտության պատմություն և մեթոդներ.
3. Բջջային տեսություն, հիմնական դրույթներ.
1. Գյուղատնտեսական արտադրության յուրահատկությունը պայմանավորված է նրանով, որ չնայած տեխնիկական գործոնների աճող դերին, կենսաբանական օբյեկտները մնում են արտադրության հիմնական գործիքներն ու միջոցները։ Ուսումնասիրության առարկաների շրջանակի և իր խորության առումով անասնաբուժությունը հետևյալն է. պաշտպանված.
Բջջաբանությունը, հյուսվածաբանությունը և սաղմնաբանությունը ֆիզիոլոգիայի, կենսաքիմիայի և այլ գիտությունների հետ միասին կազմում են ժամանակակից անասնաբուժության հիմքը։
Հյուսվածքաբանությունը (հուն. histos-հյուսվածք, logos-ուսուցում) գիտություն է կենդանիների օրգանիզմների հյուսվածքների զարգացման, կառուցվածքի և կենսագործունեության մասին։ Ժամանակակից հյուսվածաբանությունը ուսումնասիրում է կենդանիների և մարդկանց օրգանիզմների կառուցվածքները՝ կապված նրանցում տեղի ունեցող գործընթացների հետ, բացահայտում է ֆունկցիայի և կառուցվածքի միջև կապը և այլն։
Հյուսվածքաբանությունը բաժանված է 3 հիմնական բաժինների՝ բջջաբանություն կամ բջջի ուսումնասիրություն; սաղմնաբանություն կամ սաղմի ուսումնասիրություն և ընդհանուր և հատուկ հիստոլոգիա կամ հյուսվածքների, օրգանների մանրադիտակային կառուցվածքի, նրանց բջջային և հյուսվածքային կազմի ուսումնասիրություն։
Հյուսվածքաբանությունը սերտորեն կապված է մի շարք կենսաբանական և անասնաբուժական գիտությունների՝ ընդհանուր և համեմատական անատոմիայի, ֆիզիոլոգիայի, պաթոլոգիական ֆիզիոլոգիայի և պաթոլոգիական անատոմիայի, ինչպես նաև որոշ կլինիկական առարկաների հետ (ներքին բժշկություն, մանկաբարձություն և գինեկոլոգիա և այլն):
Ապագա բժիշկներին անհրաժեշտ է լավ իմացություն օրգանների բջիջների և հյուսվածքների կառուցվածքի մասին, որոնք մարմնի բոլոր տեսակի կենսագործունեության կառուցվածքային հիմքն են: Բժիշկների համար հյուսվածաբանության, բջջաբանության և սաղմնաբանության կարևորությունը նույնպես մեծանում է, քանի որ ժամանակակից անասնաբուժությունը բնութագրվում է բջջաբանական և հյուսվածքաբանական մեթոդների լայն կիրառմամբ արյան անալիզներում, ոսկրածուծի, օրգանների բիոպսիաներում և այլն:
2. Հյուսվածք հասկացությունը կենսաբանության մեջ առաջին անգամ ներմուծվել է ֆրանսիացի փայլուն երիտասարդ անատոմիստ և ֆիզիոլոգ Քսավյե Բիշատի կողմից (Bichat, 1771-1802), ով այնքան տպավորված էր տարբեր շերտերի և կառուցվածքների բազմազան հյուսվածքով, որ նա անատոմիական ուսումնասիրություններում հայտնաբերեց. նա գիրք է գրել մարմնի հյուսվածքների մասին՝ դրանում տալով դրանց ավելի քան 20 տեսակների անունները:
«Հյուսվածքաբանություն» տերմինը Բիշին չի պատկանում, չնայած նրան կարելի է համարել առաջին հյուսվածաբանը։ «Հիստոլոգիա» տերմինը Բիշայի մահից 17 տարի անց առաջարկել է գերմանացի հետազոտող Մեյերը։
Հյուսվածքը ֆիլոգենետիկորեն որոշված տարրական համակարգ է, որը միավորված է ընդհանուր կառուցվածքով, գործառույթով և զարգացմամբ (Ա.Ա. Զավարզին):
Հյուսվածքաբանության հաջողությունները սկզբնավորման պահից մինչ օրս կապված են առաջին հերթին տեխնոլոգիայի, օպտիկայի և մանրադիտակի մեթոդների զարգացման հետ։ Հյուսվածքաբանության պատմությունը կարելի է բաժանել երեք ժամանակաշրջանի՝ 1-ին` նախամիկրոսկոպիկ (մոտ 2000 տարի), 2-րդ` մանրադիտակային (մոտ 300 տարի), 3-րդ` էլեկտրոնային մանրադիտակային (մոտ 40 տարի):
Ժամանակակից հյուսվածաբանության, բջջաբանության և սաղմնաբանության մեջ օգտագործվում են հետազոտական մի շարք մեթոդներ՝ բջիջների, հյուսվածքների և օրգանների զարգացման, կառուցվածքի և ֆունկցիայի գործընթացները համակողմանիորեն ուսումնասիրելու համար:
Ուսումնասիրության առարկաները կենդանի և մեռած (ֆիքսված) բջիջներն ու հյուսվածքներն են, դրանց պատկերները՝ ստացված լուսային և էլեկտրոնային մանրադիտակներով կամ հեռուստացույցի էկրանով։ Կան մի շարք մեթոդներ, որոնք թույլ են տալիս վերլուծել այս օբյեկտները.
1) կենդանի բջիջների և հյուսվածքների ուսումնասիրման մեթոդներ. ա) մարմնի բջիջների ողջ կյանքի ընթացքում ուսումնասիրություն (in vivo) - կենդանիների մարմնում թափանցիկ խցիկներ փոխպատվաստելու մեթոդների կիրառմամբ.
բ) բջիջների և հյուսվածքների կուլտուրայի կենդանի կառուցվածքների ուսումնասիրություն (in vitro) - թերություններ. այլ բջիջների և հյուսվածքների հետ հարաբերությունները կորչում են, նյարդահումորալ կարգավորող գործոնների համալիրի ազդեցությունը և այլն.
գ) կենսական և սուպրավիտալ գունավորում, այսինքն՝ ներվիտալ գունավորում և մարմնից մեկուսացված կենդանի բջիջների ներկում:
2) մահացած բջիջների և հյուսվածքների հետազոտություն. Այստեղ ուսումնասիրության հիմնական առարկան ֆիքսված կառուցվածքներից պատրաստված հյուսվածքաբանական պատրաստուկներն են:
Լույսի և էլեկտրոնային մանրադիտակի համար հյուսվածքաբանական պատրաստման գործընթացը ներառում է հետևյալ հիմնական քայլերը. Լույսի մանրադիտակի համար անհրաժեշտ է ևս մեկ քայլ՝ հատվածները փակցնել բալասանով կամ այլ թափանցիկ միջավայրում (5):
3) բջիջների և հյուսվածքների քիմիական կազմի և նյութափոխանակության ուսումնասիրություն.
ցիտո- և հիստոքիմիական մեթոդներ,
Ռադիոավտոգրաֆիայի մեթոդ, որը հիմնված է ռադիոակտիվ տարրերի (օրինակ՝ ֆոսֆոր-32P, ածխածին -14C, ծծումբ-35S, ջրածին-3H) կամ դրա հետ պիտակավորված միացությունների օգտագործման վրա։
Դիֆերենցիալ ցենտրիֆուգացման մեթոդ - մեթոդը հիմնված է ցենտրիֆուգների օգտագործման վրա՝ րոպեում տալով 20-ից 150 հազար պտույտ: Միաժամանակ բջիջների տարբեր բաղադրամասեր առանձնացվում և նստեցվում են և որոշվում դրանց քիմիական բաղադրությունը։ - ինտերֆերոմետրիա - մեթոդը թույլ է տալիս գնահատել կենդանի և ֆիքսված բջիջներում խիտ նյութերի չոր զանգվածը և կոնցենտրացիան: - քանակական հիստոքիմիական մեթոդներ - ցիտոսպեկտրոֆոտոմետրիա - ներբջջային նյութերի քանակական ուսումնասիրության մեթոդ իրենց կլանման հատկություններով: Ցիտոսպեկտրաֆտորոմետրիան ներբջջային նյութերի ուսումնասիրման մեթոդ է՝ ըստ նրանց ֆլուորեսցենտային սպեկտրների։
4) իմունֆլյուորեսցենտային անալիզի մեթոդներ. Դրանք օգտագործվում են բջիջների դիֆերենցման գործընթացները ուսումնասիրելու, դրանցում կոնկրետ քիմիական միացություններ և կառուցվածքներ հայտնաբերելու համար։ Դրանք հիմնված են հակագեն-հակամարմին ռեակցիաների վրա:
Հիստոլոգիական պատրաստուկների մանրադիտակի մեթոդներ.
Լույսի մանրադիտակ՝ ա) ուլտրամանուշակագույն, բ) լյումինեսցենտ (լյումինեսցենտ):
Էլեկտրոնային մանրադիտակ՝ ա) փոխանցում, բ) սկանավորում (ընթերցում): Առաջինը տալիս է միայն հարթ պատկեր, երկրորդը՝ տարածական; Վերջինիս հիմնական առավելությունը (ռաստեր) դաշտի մեծ խորությունն է (100-1000 անգամ ավելի մեծ, քան լուսային մանրադիտակները), խոշորացման շարունակական փոփոխությունների լայն շրջանակը (տասնյակից մինչև տասնյակ հազարավոր անգամներ) և բարձր լուծաչափը։
3. Բարձրակարգ կենդանիների օրգանիզմը բաղկացած է մանրադիտակային տարրերից՝ բջիջներից և դրանց մի շարք ածանցյալներից՝ մանրաթելերից, ամորֆ նյութերից։
Բջջի նշանակությունը բազմաբջիջ օրգանիզմում որոշվում է նրանով, որ դրա միջոցով փոխանցվում է ժառանգական ինֆորմացիա, դրանով է սկսվում բազմաբջիջ կենդանիների զարգացումը. Բջիջների գործունեության շնորհիվ ձևավորվում են ոչ բջջային կառուցվածքներ և հիմնական նյութ, որոնք բջիջների հետ միասին կազմում են բարդ օրգանիզմում հատուկ գործառույթներ կատարող հյուսվածքներ և օրգաններ։ Դյուտրոշեն (1824, 1837) և Շվանը (1839) պետք է համարել բջջային տեսության ստեղծող։
Դյուտրոշետ (1776-1847) - կենդանաբան, բուսաբան, մորֆոլոգ, ֆիզիոլոգ։ 1824 թվականին նա հրատարակեց իր «Անատոմիական և ֆիզիոլոգիական հետազոտություններ կենդանիների և բույսերի նուրբ կառուցվածքի և նրանց շարժունակության մասին» գիրքը։
Բջջային տեսության ստեղծմանը նախորդել են հետեւյալ բացահայտումները. 1610 թվականին 46-ամյա պրոֆ. Պադուայի համալսարանի մաթեմատիկոս Գ. Գալիլեոն նախագծել է մանրադիտակ: 1665 թվականին Ռոբերտ Հուկը հայտնաբերեց բջիջը 100 անգամ մեծացմամբ։ Նրա ժամանակակիցը՝ Ֆելիս Ֆոնտանան, ասել է. Հուկի «Միկրոգրաֆիան» ներառում էր 54 դիտարկում, այդ թվում՝ «Դիտարկում 18. Խցանափայտի սխեմայի կամ կառուցվածքի կամ որոշ այլ չամրացված մարմինների բջիջների և ծակոտիների վրա»։
Պատմությունից. 1645 թվականին Լոնդոնում ապրող երիտասարդների (ուսանողների) մի խումբ դասերից հետո սկսեց ամեն օր հավաքվել՝ քննարկելու փորձարարական փիլիսոփայության խնդիրները։ Նրանց թվում էին Ռոբերտ Բոյլը (18 տարեկան), Ռ. Հուկը (17 տարեկան), Ռենը (23 տարեկան) և այլք։Այսպես ծնվեց Բրիտանական ակադեմիան, այնուհետև Լոնդոնի թագավորական ընկերությունը (Չարլզ II-ը նրա պատվին էր։ անդամ).
Կենդանական բջիջը հայտնաբերել է Անտոն վան Լեուվենհուկը (1673-1695): Նա ապրում էր Դելֆտում և գործի առևտուր էր անում։ Նա իր մանրադիտակները հասցրեց մինչև 275 x: Պետրոս I-ը ցույց տվեց արյան շրջանառություն օձաձկան թրթուրի պոչում:
Ներկայումս բջջային տեսությունը ասում է. 1) բջիջը կենդանի էակի ամենափոքր միավորն է, 2) տարբեր օրգանիզմների բջիջները կառուցվածքով նման են, 3) բջիջների վերարտադրությունը տեղի է ունենում սկզբնական բջիջը բաժանելով, 4) բազմաբջիջ օրգանիզմները բարդ անսամբլներ են։ բջիջների և դրանց ածանցյալների՝ համակցված հյուսվածքների և օրգանների ամբողջական ինտեգրված համակարգերի մեջ, որոնք ենթակա և փոխկապակցված են կարգավորման միջբջջային, հումորալ և նյարդային ձևերով։
Բջջ - կյանքի տարրական միավոր
1. Կենդանի նյութի կազմը և ֆիզիկաքիմիական հատկությունները.
2. Բջիջների տեսակները. Էուկարիոտիկ բջջի ծագման տեսություններ.
3. Բջջային թաղանթները, դրանց մոլեկուլային կազմը և գործառույթները:
1. Տիպիկ բջիջը միջուկով, ցիտոպլազմայով և դրանում պարունակվող բոլոր օրգանելներով դեռ չի կարող համարվել կենդանի նյութի կամ պրոտոպլազմայի ամենափոքր միավորը (հունարեն «պրոտոս»՝ առաջին, «պլազմա»՝ կրթություն): Կան նաև կյանքի ավելի պարզունակ կամ ավելի պարզ կազմակերպված միավորներ՝ այսպես կոչված պրոկարիոտային օրգանիզմներ (հունարեն «կարիոն»՝ միջուկ), որոնք ներառում են վիրուսների, բակտերիաների և որոշ ջրիմուռների մեծ մասը; նրանք, ի տարբերություն իրական միջուկով ավելի բարձր տեսակի բջիջների (էուկարիոտիկ բջիջներ), չունեն միջուկային թաղանթ, և միջուկային նյութը խառնվում է կամ ուղղակիորեն շփվում է պրոտոպլազմայի մնացած մասի հետ:
Կենդանի նյութի կազմը ներառում է սպիտակուցներ, նուկլեինաթթուներ (ԴՆԹ և ՌՆԹ), պոլիսախարիդներ և լիպիդներ։ Բջջի քիմիական բաղադրիչները կարելի է բաժանել անօրգանական (ջուր և հանքային աղեր) և օրգանական (սպիտակուցներ, ածխաջրեր, նուկլեինաթթուներ, լիպիդներ և այլն):
Բուսական և կենդանական բջիջների ցիտոպլազմը պարունակում է 75-85% ջուր, 10-20% սպիտակուց, 2-3% լիպիդներ, 1% ածխաջրեր և 1% ոչ: օրգանական նյութեր.
ԴՆԹ-ն մոլեկուլ է (այն պարունակում է 0,4%), որը պարունակում է գենետիկ տեղեկատվություն, որն ուղղորդում է հատուկ բջջային սպիտակուցների սինթեզը։ Մեկ ԴՆԹ-ի մոլեկուլում կա մոտ 44 ՌՆԹ մոլեկուլ, 700 սպիտակուցի մոլեկուլ և 7000 լիպիդային մոլեկուլ։
ՌՆԹ-ի առաջնային կառուցվածքը նման է ԴՆԹ-ին, միայն թե ՌՆԹ-ն տիմինի փոխարեն պարունակում է ռիբոզ և ուրացիլ։ Այժմ հաստատվել է, որ կան երեք տեսակի ՌՆԹ, որոնք տարբերվում են մոլեկուլային քաշով և այլ հատկություններով՝ ռիբոսոմային, տեղեկատվական և տրանսպորտային։ ՌՆԹ-ի այս երեք տեսակները սինթեզվում են միջուկում և մասնակցում են սպիտակուցի սինթեզին։
2. Shutton (1925) բոլոր կենդանի օրգանիզմները բաժանել է երկու տեսակի (կլիստերի)՝ պրոկարիոտների և էուկարիոտների։ Նրանք շեղվել են նախաքեմբրյան շրջանում (600-4500 միլիոն տարի առաջ)։ Էուկարիոտիկ բջջի ծագման երկու հասկացություն կա՝ էկզոգեն (սիմբիոտիկ) և էնդոգեն։ Առաջինը հիմնված է տարբեր պրոկարիոտ օրգանիզմների միմյանց հետ համակցելու սկզբունքի ճանաչման վրա։ Էնդոգեն հայեցակարգը հիմնված է ուղղակի ֆիլիացիայի սկզբունքի վրա, այսինքն. պրոկարիոտ օրգանիզմների հաջորդական էվոլյուցիոն փոխակերպումը էուկարիոտների։
Կաթնասունների օրգանիզմում հյուսվածաբանները թվարկում են մոտ 150 տեսակի բջիջներ, և դրանցից շատերը հարմարեցված են մեկ կոնկրետ առաջադրանք կատարելու համար։ Բջջի ձևն ու կառուցվածքը կախված է նրա կատարած գործառույթից:
Բջիջների գործառույթները՝ դյուրագրգռություն, կծկողականություն, սեկրեցիա, շնչառություն, անցում, ներծծում և յուրացում, արտազատում, աճ և վերարտադրություն:
3. Ցանկացած բջիջ սահմանազատված է պլազմային թաղանթով: Այն այնքան բարակ է, որ չի երևում լուսային մանրադիտակի տակ։ Պլազմային թաղանթը, որը հեշտությամբ վնասվում է միկրոասեղով, կարող է վերականգնվել, բայց ավելի ծանր վնասով, հատկապես կալցիումի իոնների բացակայության դեպքում, ցիտոպլազմը դուրս է հոսում պունկցիայի միջոցով և բջիջը մահանում է:
Համաձայն ժամանակակից տեսություն, պլազմային թաղանթը բաղկացած է բևեռային լիպիդների երկշերտից և դրա մեջ ներկառուցված գնդային սպիտակուցի մոլեկուլներից։ Այս շերտերի շնորհիվ թաղանթն ունի առաձգականություն և հարաբերական մեխանիկական ուժ։ Բջջային տեսակների մեծ մասի պլազմային թաղանթը բաղկացած է երեք շերտից՝ յուրաքանչյուրը մոտավորապես 2,5 նմ լայնությամբ: Նմանատիպ կառուցվածք, որը կոչվում է «տարրական թաղանթ», հայտնաբերված է նաև ներբջջային թաղանթների մեծ մասում։ Կենսաքիմիական անալիզը ցույց է տվել, որ դրանցում լիպիդներն ու սպիտակուցները պարունակվում են 1,0:1,7 հարաբերակցությամբ։ Սպիտակուցի բաղադրիչը, որը կոչվում է ստրոմատին, թթվային մանրաթելային սպիտակուց է՝ բարձր մոլեկուլային քաշով: Լիպիդային բաղադրիչների հիմնական մասը ձևավորվում է ֆոսֆոլիպիդներով, հիմնականում լեցիտինով և ցեֆալինով:
Պլազմալեման բջջային թաղանթ է, որն իրականացնում է սահմանազատող, փոխադրող և ընկալիչի գործառույթներ: Այն ապահովում է բջիջների և միջբջջային փոխազդեցությունների մեխանիկական հաղորդակցություն, պարունակում է բջջային ընկալիչներ հորմոնների և բջիջը շրջապատող միջավայրի այլ ազդանշանների համար, նյութերը բջջից տեղափոխում է բջջ՝ ինչպես կոնցենտրացիայի գրադիենտի երկայնքով՝ պասիվ փոխանցում, այնպես էլ էներգիայի ծախսերով՝ ընդդեմ կոնցենտրացիայի գրադիենտի։ - ակտիվ փոխանցում:
Մեմբրանը բաղկացած է պլազմային թաղանթից, ոչ թաղանթային համալիրից՝ գլիկոկալեքսից և ենթամեմբրանային հենաշարժական ապարատից։
Գլիկոկալեքսը պարունակում է մոտ 1% ածխաջրեր, որոնց մոլեկուլները կազմում են պոլիսախարիդների երկար ճյուղավորված շղթաներ՝ կապված թաղանթային սպիտակուցների հետ։ Գլիկոկալեքսում տեղակայված սպիտակուցները՝ ֆերմենտները մասնակցում են նյութերի վերջնական արտաբջջային տրոհմանը: Այս ռեակցիաների արգասիքները մոնոմերների տեսքով մտնում են բջիջ։ Ակտիվ տեղափոխմամբ նյութերի տեղափոխումը բջիջ իրականացվում է կա՛մ մոլեկուլների ներթափանցմամբ լուծույթի տեսքով՝ պինոցիտոզով, կա՛մ խոշոր մասնիկների գրավմամբ՝ ֆագոցիտոզով։
Հյուսվածքների ֆունկցիոնալ և մորֆոլոգիական առանձնահատկություններին համապատասխան, բջջային թաղանթը կազմում է նրանց բնորոշ միջբջջային շփումների ապարատը։ Նրանց հիմնական ձևերն են՝ պարզ կոնտակտային (կամ կպչուն գոտի), ամուր (փակվող) և բաց կոնտակտ։ Դեսմոսոմները ամուր շփման տեսակ են:
Կենսաբանական թաղանթները գործում են որպես ցրված խոչընդոտներ: K+, Na+, Cl- և այլն իոնների, ինչպես նաև մակրոմոլեկուլային միացությունների ընտրովի թափանցելիության շնորհիվ նրանք սահմանազատում են ներբջջային և միջբջջային ռեակցիայի գոտիները և ստեղծում էլեկտրական և նյութերի կոնցենտրացիայի գրադիենտներ։ Սա հնարավոր է դարձնում հատուկ գործառույթներով պատվիրված կենսաբանական կառույցների առկայությունը։
Նյութերի մուտքը բջիջ կոչվում է էնդոցիտոզ: Բայց կա նաև էկզոցիտոզ: Օրինակ, արտազատվող վեզիկուլները փակվում են Գոլջիի ապարատից՝ գաղթելով դեպի բջջային թաղանթ և դուրս նետելով դրանց պարունակությունը։ Այս դեպքում վեզիկուլային թաղանթը միաձուլվում է իրեն հոմոլոգ բջջաթաղանթի հետ։
Էլեկտրոնային մանրադիտակային տվյալների հիման վրա կարելի է ենթադրել, որ պլազմային թաղանթը Գոլջիի ապարատի արտադրանք է։ Այս օրգանելից, շարունակաբար բաժանվող վեզիկուլների տեսքով, տեղի է ունենում թաղանթային նյութի մշտական տեղափոխում («մեմբրանային հոսք»), որը վերականգնում է պլազմոլեմայի օգտագործված հատվածները և ապահովում դրա աճը բջիջների բաժանումից հետո։
Մեմբրանը տեսակների հատուկ և բջիջներին հատուկ մակերևութային հատկությունների կրող է, կապված դրա վրա գլիկոզամինոգլիկանների և սպիտակուցների բնորոշ բաշխման հետ: Նրանց մոլեկուլները կարող են նաև ծածկել բջիջների մակերեսը ամենաբարակ թաղանթների տեսքով և միջբջջային մատրիցա կազմել հարևան բջիջների միջև։ Բջջային շփման հատկությունները և իմունային պատասխանները որոշվում են այս մեմբրանի բաղադրիչներով:
Բազմաթիվ բջիջներ, հատկապես կլանման համար մասնագիտացված (աղիքային էպիթելիա), ունեն մազի նման աճեր՝ արտաքին կողմում միկրովիլիներ: Ձևավորված կամ «խոզանակի եզրագիծը» կրում է ֆերմենտներ, մասնակցում է նյութերի քայքայմանը և փոխադրման գործընթացներին: Հեղուկի ինտենսիվ անցման համար մասնագիտացված բջիջների բազալային կողմում (օսմոկարգավորման ժամանակ), օրինակ, երիկամային խողովակների և մալպիգիական անոթների էպիթելիում, թաղանթը ձևավորում է բազմաթիվ ինվագինացիաներ, որոնք կազմում են բազալ լաբիրինթոսը: Բջջային սեկրեցիայի արգասիքը՝ նկուղային թաղանթը, հաճախ բաժանում է էպիթելը ավելի խորը բջիջների շերտերից։
Հարևան բջիջների շփման կետերում առաջանում են հատուկ թաղանթային կառուցվածքներ։ Կան տարածքներ, որտեղ թաղանթները այնքան մոտ են միմյանց, որ միջբջջային նյութի համար տեղ չկա (սեղմ շփում): Մյուս տարածքներում հայտնվում են բարդ կոնտակտային օրգանելներ՝ դեզմոսոմներ։ Նրանք և այլ կոնտակտային կառույցներ ծառայում են մեխանիկական միացման համար և, ամենակարևորը, ապահովում են հարևան բջիջների քիմիական և էլեկտրական ինտեգրումը, ինչը հեշտացնում է միջբջջային իոնների տեղափոխումը ցածր էլեկտրական դիմադրության պատճառով:
Կենդանական բջիջի կառուցվածքը
1. Ցիտոպլազմա և օրգանելներ, նրանց գործառույթը:
2. Միջուկը, նրա կառուցվածքը և գործառույթները:
3. Բաժանման տեսակները, բջջային ցիկլի փուլերը.
1. Շրջակա միջավայրից պլազմոլեմայով անջատված ցիտոպլազմը ներառում է հիալոպլազմը, նրանում առկա պարտադիր բջջային բաղադրիչները՝ օրգանելները, ինչպես նաև տարբեր ոչ մշտական կառուցվածքներ՝ ներդիրներ (նկ. 1)։
Hyaloplasm (hyalinos - թափանցիկ) - ցիտոպլազմայի հիմնական պլազման կամ մատրիցը, բջջի շատ կարևոր մասն է, նրա իրական ներքին միջավայրը:
Էլեկտրոնային մանրադիտակում մատրիցը նման է միատարր և մանրահատիկ նյութի՝ ցածր էլեկտրոնային խտությամբ: Հիալոպլազմը բարդ կոլոիդային համակարգ է, որը ներառում է տարբեր կենսապոլիմերներ՝ սպիտակուցներ, նուկլեինաթթուներ, պոլիսախարիդներ և այլն։ Հիալոպլազմայի կազմը հիմնականում բաղկացած է տարբեր գնդաձեւ սպիտակուցներից։ Նրանք կազմում են էուկարիոտ բջջի ընդհանուր սպիտակուցի 20-25%-ը։ Հիալոպլազմայի ամենակարևոր ֆերմենտները ներառում են շաքարների, ազոտային հիմքերի, ամինաթթուների, լիպիդների և այլ կարևոր միացությունների նյութափոխանակության ֆերմենտներ: Հիալոպլազմում առկա են սպիտակուցների սինթեզում ամինաթթուների ակտիվացման ֆերմենտներ, տրանսպորտային ՌՆԹ (tRNA): Հիալոպլազմում, ռիբոսոմների և պոլիռիբոսոմների մասնակցությամբ, տեղի է ունենում սպիտակուցների սինթեզ, որոնք անհրաժեշտ են բջջի պատշաճ կարիքների համար՝ այս բջջի կյանքը պահպանելու և ապահովելու համար։
Օրգանելները միկրոկառուցվածքներ են, որոնք մշտապես առկա են և պարտադիր են բոլոր բջիջների համար՝ կատարելով կենսական գործառույթներ։
Կան թաղանթային օրգանելներ՝ միտոքոնդրիաներ, էնդոպլազմային ցանց (հատիկավոր և հարթ), Գոլջիի ապարատ, լիզոսոմներ, իսկ պլազմոլեմման նույնպես պատկանում է թաղանթային օրգանելների կատեգորիային. ոչ թաղանթային օրգանելներ՝ ազատ ռիբոսոմներ և պոլիսոմներ, միկրոխողովակներ, ցենտրիոլներ և թելեր (միկրաթելեր): Շատ բջիջներում օրգանելները կարող են մասնակցել մասնագիտացված բջիջներին բնորոշ հատուկ կառուցվածքների ձևավորմանը։ Այսպիսով, թարթիչները և դրոշակները ձևավորվում են ցենտրիոլների և պլազմային թաղանթի շնորհիվ, միկրովիլիները պլազմային մեմբրանի ելքեր են հիալոպլազմով և միկրոթելերով, սերմնաբջիջների ակրոսոմը Գոլջիի ապարատի տարրերի ածանցյալն է և այլն:
Նկ 1. Կենդանական օրգանիզմների բջջի ուլտրամիկրոսկոպիկ կառուցվածքը (սխեմա)
1 - միջուկ; 2 - պլազմալեմա; 3 - միկրովիլի; 4 - ագրարային էնդոպլազմիկ ցանց; 5 - հատիկավոր էնդոպլազմիկ ցանց; 6 - Golgi ապարատ; 7 - ցենտրիոլ և բջջային կենտրոնի միկրոխողովակներ; 8 - միտոքոնդրիա; 9 - ցիտոպլազմիկ վեզիկուլներ; 10 - լիզոսոմներ; 11 - միկրոթելեր; 12 - ռիբոսոմներ; 13 - սեկրեցիայի հատիկների մեկուսացում.
Մեմբրանի օրգանելները ցիտոպլազմայի միայնակ կամ փոխկապակցված բաժանմունքներ են, որոնք սահմանազատված են շրջապատող հիալոպլազմից թաղանթով և ունեն իրենց բովանդակությունը՝ տարբեր կազմով, հատկություններով և գործառույթներով.
Միտոքոնդրիաները ATP-ի սինթեզի օրգանելներն են: Նրանց հիմնական գործառույթը կապված է օրգանական միացությունների օքսիդացման և այդ միացությունների քայքայման ժամանակ արձակված էներգիայի օգտագործման հետ՝ ATP մոլեկուլների սինթեզի համար։ Միտոքոնդրիաները կոչվում են նաև բջջի էներգետիկ կայաններ կամ բջջային շնչառության օրգանելներ։
«Միտոխոնդրիոն» տերմինը ներկայացվել է Բենդան 1897 թվականին։ Կենդանի բջիջներում կարելի է նկատել միտոքոնդրիաներ, քանի որ ունեն բավականին բարձր խտություն։ Կենդանի բջիջներում միտոքոնդրիումները կարող են շարժվել, միաձուլվել միմյանց հետ և բաժանվել։ Կենդանական բջիջների միտոքոնդրիաների ձևն ու չափը բազմազան են, բայց միջինում դրանց հաստությունը կազմում է մոտ 0,5 մկմ, իսկ երկարությունը՝ 1-ից մինչև 10 մկմ։ Նրանց թիվը բջիջներում շատ է տատանվում՝ առանձին տարրերից մինչև հարյուրավոր: Այսպիսով, լյարդի բջիջում նրանք կազմում են ցիտոպլազմայի ընդհանուր ծավալի ավելի քան 20% -ը: Լյարդի բջջի բոլոր միտոքոնդրիաների մակերեսը 4-5 անգամ ավելի մեծ է, քան նրա պլազմային մեմբրանի մակերեսը:
Միտոքոնդրիաները սահմանափակված են մոտ 7 նմ հաստությամբ երկու թաղանթով։ Արտաքին միտոքոնդրիալ թաղանթը սահմանափակում է միտոքոնդրիոնի իրական ներքին պարունակությունը, նրա մատրիցը: բնորոշ հատկանիշՄիտոքոնդրիայի ներքին թաղանթները միտոքոնդրիայի ներսում բազմաթիվ ելուստներ ստեղծելու նրանց ունակությունն է: Նման ինվագինացիաները հաճախ նման են հարթ սրածայրերի կամ կրիստաների: Միտոքոնդրիալ մատրիցային շղթաները ԴՆԹ մոլեկուլներ են, իսկ փոքր հատիկները՝ միտոքոնդրիալ ռիբոսոմներ։
Էնդոպլազմիկ ցանցը հայտնաբերել է Կ.Ռ. Փորթերը 1945թ.-ին: Այս օրգանիլը վակուոլների, հարթ թաղանթային պարկերի կամ խողովակային գոյացությունների հավաքածու է, որոնք ստեղծում են, ասես, թաղանթային ցանց ցիտոպլազմայի ներսում: Կան երկու տեսակ՝ հատիկավոր և հարթ էնդոպլազմիկ ցանց։
Հատիկավոր էնդոպլազմիկ ցանցը ներկայացված է փակ թաղանթներով, որոնց տարբերակիչ առանձնահատկությունն այն է, որ դրանք հիալոպլազմայի կողմից պատված են ռիբոսոմներով։ Ռիբոսոմները ներգրավված են տվյալ բջջից ստացված սպիտակուցների սինթեզում։ Բացի այդ, հատիկավոր էնդոպլազմիկ ցանցը ներգրավված է ներբջջային նյութափոխանակության կազմակերպման համար անհրաժեշտ ֆերմենտային սպիտակուցների սինթեզում, ինչպես նաև օգտագործվում է ներբջջային մարսողության համար։
Ցանցի խոռոչներում կուտակված սպիտակուցները, շրջանցելով հիալոպլազմը, կարող են տեղափոխվել Գոլջիի համալիրի վակուոլներ, որտեղ դրանք հաճախ ձևափոխվում են և կազմում են լիզոսոմների կամ սեկրետորային հատիկներ:
Հատիկավոր էնդոպլազմիկ ցանցի դերը կայանում է նրանում, որ արտահանվող սպիտակուցները սինթեզում են նրա պոլիսոմների վրա, թաղանթային խոռոչների ներսում հիալոպլազմայի պարունակությունից դրանց մեկուսացման, այդ սպիտակուցների տեղափոխման մեջ բջջի այլ մասեր և սինթեզում: բջջային թաղանթների կառուցվածքային բաղադրիչները.
Ագրանուլյար (հարթ) էնդոպլազմիկ ցանցը նույնպես ներկայացված է թաղանթներով, որոնք կազմում են փոքր վակուոլներ և խողովակներ, խողովակներ, որոնք կարող են ճյուղավորվել միմյանց հետ։ Ի տարբերություն հատիկավոր ER-ի, հարթ ER-ի թաղանթների վրա ռիբոսոմներ չկան: Վակուոլների և խողովակների տրամագիծը սովորաբար կազմում է մոտ 50-100 նմ։
Հարթ էնդոպլազմիկ ցանցը առաջանում և զարգանում է հատիկավոր էնդոպլազմիկ ցանցի հաշվին։
Հարթ EPS-ի ակտիվությունը կապված է լիպիդների և որոշ ներբջջային պոլիսախարիդների նյութափոխանակության հետ։ Smooth ER-ը մասնակցում է լիպիդների սինթեզի վերջին փուլերին: Այն բարձր զարգացած է մակերիկամի կեղևի և ամորձու սուստենտոցիտների (Սերտոլի բջիջներ) ստերոիդներ արտազատող բջիջներում:
Զոլավոր մկանային մանրաթելերում հարթ ER-ն ի վիճակի է կուտակել կալցիումի իոնները, որոնք անհրաժեշտ են մկանային հյուսվածքի աշխատանքի համար:
Շատ կարևոր է հարթ EPS-ի դերը օրգանիզմի համար վնասակար տարբեր նյութերի ապաակտիվացման գործում։
Golgi համալիր (CG). 1898թ.-ին Կ.Գոլջին, օգտագործելով ծանր մետաղները բջջային կառուցվածքներին կապելու հատկությունները, հայտնաբերեց ցանցային գոյացություններ նյարդային բջիջներում, որոնք նա անվանեց ներքին ցանցային ապարատ:
Այն ներկայացված է փոքր գոտում միասին հավաքված թաղանթային կառուցվածքներով։ Այս թաղանթների կուտակման առանձին գոտին կոչվում է դիկտյոսոմ։ Բջիջում կարող են լինել մի քանի նման գոտիներ։ Դիկտոսոմում իրար մոտ տեղակայված են 5-10 հարթ ցիստեռններ (20-25 նմ հեռավորության վրա), որոնց միջև կան հիալոպլազմայի բարակ շերտեր։ Բացի ցիստեռններից, CG գոտում նկատվում են բազմաթիվ փոքր վեզիկուլներ (վեզիկուլներ): CG-ն մասնակցում է ցիտոպլազմային ցանցում սինթեզված արտադրանքների տարանջատմանը և կուտակմանը, դրանց քիմիական վերադասավորումներին, հասունացմանը. CG-ի ցիստեռններում տեղի է ունենում պոլիսախարիդների սինթեզ, դրանց կոմպլեքսավորումը սպիտակուցներով և, որ ամենակարևորն է, բջջից դուրս պատրաստի գաղտնիքների հեռացում։
Լիզոսոմները 0,2-0,4 մկմ չափերով գնդաձեւ կառուցվածքների բազմազան դաս են, որոնք սահմանափակված են մեկ թաղանթով:
Լիզոսոմների բնորոշ առանձնահատկությունը նրանց մեջ հիդրոլիտիկ ֆերմենտների առկայությունն է, որոնք քայքայում են տարբեր կենսապոլիմերներ։ Լիզոսոմները հայտնաբերվել են 1949 թվականին դե Դյուվի կողմից։
Պերօքսիզոմները փոքր 0,3-1,5 մկմ օվալաձև մարմիններ են, որոնք սահմանափակված են թաղանթով: Դրանք հատկապես բնորոշ են լյարդի և երիկամների բջիջներին։ Ամինաթթուների օքսիդացման ֆերմենտները առաջացնում են ջրածնի պերօքսիդ, որը քայքայվում է կատալազ ֆերմենտի կողմից: Պերօքսիզոմային կատալազը կարևոր պաշտպանիչ դեր է խաղում, քանի որ H2O2-ը թունավոր նյութ է բջջի համար:
Ոչ թաղանթային օրգանելներ
Ռիբոսոմները՝ սպիտակուցի, պոլիպեպտիդային մոլեկուլների սինթեզի տարրական ապարատը, հանդիպում են բոլոր բջիջներում: Ռիբոսոմները բարդ ռիբոնուկլեոպրոտեիններ են, որոնք ներառում են սպիտակուցներ և ՌՆԹ մոլեկուլներ։ Էուկարիոտիկ բջիջների գործող ռիբոսոմի չափը 25 x 20 x 20 նմ է։
Կան միայնակ ռիբոսոմներ և բարդ ռիբոսոմներ (պոլիսոմներ): Ռիբոսոմները կարող են ազատ տեղակայվել հիալոպլազմայում և կապված լինել էնդոպլազմային ցանցի թաղանթների հետ։ Ազատ ռիբոսոմները սպիտակուցներ են կազմում հիմնականում բջջի սեփական կարիքների համար, կապված ապահովում են սպիտակուցների սինթեզ «արտահանման համար»։
Microtubules-ը սպիտակուցային բնույթի ֆիբրիլային բաղադրիչներ են: Ցիտոպլազմայում նրանք կարող են ձևավորել ժամանակավոր գոյացություններ (spindle): Միկրոխողովակները ցենտրիոլների մի մասն են և հանդիսանում են նաև թարթիչների և դրոշակների հիմնական կառուցվածքային տարրերը: Դրանք ուղիղ, չճյուղավորված երկար խոռոչ գլաններ են։ Նրանց արտաքին տրամագիծը մոտ 24 նմ է, ներքին լույսը՝ 15 նմ, իսկ ցանցի հաստությունը՝ 5 նմ։ Միկրոխողովակները պարունակում են տուբուլիններ կոչվող սպիտակուցներ: Ստեղծելով ներբջջային կմախք՝ միկրոխողովակները կարող են լինել բջջի և նրա ներբջջային բաղադրիչների կողմնորոշված շարժման գործոններ՝ ստեղծելով տարբեր նյութերի ուղղորդված հոսքերի գործոններ:
Ցենտրիոլներ. Տերմինը առաջարկվել է Տ. Բովերիի կողմից 1895 թվականին՝ վերաբերելու շատ փոքր մարմիններին։ Ցենտրիոլները սովորաբար դասավորված են զույգով՝ դիպլոսոմով, շրջապատված են ավելի թեթև ցիտոպլազմայի գոտիով, որից տարածվում են շառավղային բարակ մանրաթելեր (ցենտոսֆերա)։ Ցենտրիոլների և ցենտոսֆերայի համակցությունը կոչվում է բջջի կենտրոն։ Այս օրգանելները բաժանվող բջիջներում մասնակցում են բաժանման spindle-ի ձևավորմանը և գտնվում են նրա բևեռներում: Չբաժանվող բջիջներում դրանք գտնվում են ԿԳ-ի մոտ։
Ցենտրիոլների կառուցվածքի հիմքում ընկած են 9 եռյակ միկրոխողովակները, որոնք տեղակայված են շրջագծի շուրջ, այդպիսով ձևավորելով խոռոչ գլան: Նրա լայնությունը մոտ 0,2 միկրոն է, իսկ երկարությունը՝ 0,3-0,5 մկմ։
Բացի միկրոխողովակներից, ցենտրիոլները ներառում են լրացուցիչ կառուցվածքներ՝ եռյակներ միացնող «բռնակներ»: Ցենտրիոլային միկրոխողովակային համակարգերը կարելի է նկարագրել բանաձևով. (9 x 3) + 0, ընդգծելով դրա կենտրոնական մասում միկրոխողովակների բացակայությունը:
Բջիջները միտոտիկ բաժանման նախապատրաստելիս տեղի է ունենում ցենտրիոլների կրկնապատկում:
Ենթադրվում է, որ ցենտրիոլները ներգրավված են տուբուլինի կողմից պոլիմերացման ինդուկցիայի մեջ միկրոխողովակների ձևավորման ժամանակ: Միտոզից առաջ ցենտրիոլը հանդիսանում է բջիջների բաժանման spindle microtubules-ի պոլիմերացման կենտրոններից մեկը։
Կիլիա և դրոշակ: Սրանք շարժման հատուկ օրգանելներ են։ Ցիտոպլազմայի թարթիչների և դրոշակի հիմքում տեսանելի են փոքր հատիկներ՝ բազալային մարմիններ։ Թարթիչների երկարությունը 5-10 միկրոն է, դրոշակները՝ մինչև 150 միկրոն։
Թարթիչը ցիտոպլազմայի բարակ գլանաձեւ ելքն է՝ 200 նմ տրամագծով։ Այն պատված է պլազմային թաղանթով։ Ներսում կա աքսոնեմ («առանցքային թել»), որը բաղկացած է միկրոխողովակներից։
Աքսոնեմը պարունակում է 9 կրկնակի միկրոխողովակներ: Այստեղ միկրոխողովակների համակարգը շրջապատված է թարթիչներով (9 x 2) + 2:
Թարթիչներով և դրոշակներով ազատ բջիջները շարժվելու հատկություն ունեն։ Նրանց շարժման ձևը «սահող թելեր» է։
Ցիտոպլազմայի մանրաթելային բաղադրիչները ներառում են 5-7 նմ հաստությամբ միկրոթելեր և, այսպես կոչված, միջանկյալ թելեր՝ միկրոֆիբրիլներ՝ մոտ 10 նմ հաստությամբ։
Միկրաթելերը հայտնաբերված են բոլոր տեսակի բջիջներում: Կառուցվածքով և գործառույթով դրանք տարբեր են, բայց մորֆոլոգիապես դժվար է տարբերակել միմյանցից։ Նրանց քիմիական բաղադրությունը տարբեր է. Նրանք կարող են կատարել ցիտոկմախքի գործառույթները և մասնակցել բջջի ներսում շարժման ապահովմանը:
Միջանկյալ թելերը նույնպես սպիտակուցային կառուցվածքներ են։ Էպիթելիում դրանք ներառում են կերատին: Թելերի կապոցները ձեւավորում են տոնոֆիբրիլներ, որոնք տեղավորվում են դեզմոսոմների մեջ։ Միջանկյալ միկրոթելերի դերը, ամենայն հավանականությամբ, աջակցության շրջանակ է:
ընդգրկումներ ցիտոպլազմայում. Սրանք բջջի կամընտիր բաղադրիչներն են, որոնք առաջանում և անհետանում են՝ կախված բջիջների նյութափոխանակության վիճակից: Տարբերում են տրոֆիկ, արտազատող, արտազատող և պիգմենտային ներդիրներ։ Տրոֆիկ ներդիրները չեզոք ճարպեր և գլիկոգեն են: Պիգմենտային ներդիրները կարող են լինել էկզոգեն (կարոտին, ներկանյութեր, փոշու մասնիկներ և այլն) և էնդոգեն (հեմոգլոբին, մելանին և այլն)։ Նրանց առկայությունը ցիտոպլազմայում կարող է փոխել հյուսվածքի գույնը: Հաճախ հյուսվածքների պիգմենտացիան ծառայում է որպես ախտորոշիչ նշան:
Միջուկն ապահովում է ընդհանուր գործառույթների երկու խումբ՝ մեկը կապված է գենետիկական տեղեկատվության իրական պահպանման և փոխանցման հետ, մյուսը՝ դրա իրականացման, սպիտակուցի սինթեզի ապահովման հետ։
ԴՆԹ-ի մոլեկուլների վերարտադրումը կամ վերարտադրումը տեղի է ունենում միջուկում, ինչը հնարավորություն է տալիս երկու դուստր բջիջներին միտոզի ընթացքում ստանալ գենետիկական տեղեկատվության ճիշտ նույն քանակական և որակական ծավալները:
Միջուկի ակտիվությամբ ապահովված բջջային պրոցեսների մեկ այլ խումբ սպիտակուցի սինթեզի սեփական ապարատի ստեղծումն է։ Սա ոչ միայն տարբեր սուրհանդակային ՌՆԹ-ների ԴՆԹ մոլեկուլների սինթեզն է, տրանսկրիպցիան, այլ նաև բոլոր տեսակի տրանսպորտային և ռիբոսոմային ՌՆԹ-ների տրանսկրիպցիան:
Այսպիսով, միջուկը ոչ միայն գենետիկական նյութի ընդունարան է, այլև այն վայր, որտեղ այդ նյութը գործում և վերարտադրվում է:
Չբաժանվող, միջֆազային բջիջի միջուկը սովորաբար մեկ բջջի մեկ է: Միջուկը բաղկացած է քրոմատինից, միջուկից, կարիոպլազմայից (նուկլեոպլազմա) և միջուկային ծրարից, որը բաժանում է այն ցիտոպլազմայից (կարիոլեմմա)։
Կարիոպլազմը կամ միջուկային հյութը միջուկի միկրոսկոպիկ կառուցվածք չունեցող նյութ է։ Այն պարունակում է տարբեր սպիտակուցներ (նուկլեոպրոտեիններ, գլիկոպրոտեիններ), ֆերմենտներ և միացություններ, որոնք մասնակցում են նուկլեինաթթուների, սպիտակուցների և կարիոպլազմը կազմող այլ նյութերի սինթեզին։ Էլեկտրոն - միջուկային հյութում միկրոսկոպիկ կերպով հայտնաբերվում են 15 նմ տրամագծով ռիբոնուկլեոպրոտեինի հատիկներ:
Միջուկային հյութում հայտնաբերվել են նաև գլիկոլիտիկ ֆերմենտներ, որոնք մասնակցում են ազատ նուկլեոտիդների և դրանց բաղադրիչների սինթեզին և տրոհմանը, սպիտակուցի և ամինաթթուների նյութափոխանակության ֆերմենտներ: Միջուկի կենսագործունեության բարդ պրոցեսներն ապահովվում են գլիկոլիզի գործընթացում արտազատվող էներգիայով, որի ֆերմենտները պարունակվում են միջուկային հյութում։
Քրոմատին. Քրոմատինը պարունակում է ԴՆԹ՝ սպիտակուցի հետ միասին։ Քրոմոսոմները, որոնք հստակ տեսանելի են միտոտիկ բջիջների բաժանման ժամանակ, ունեն նույն հատկությունները։ Միջֆազային միջուկների քրոմատինը քրոմոսոմ է, որն այս պահին կորցնում է իր կոմպակտ ձևը, թուլանում, խտանում է։ Ամբողջական խտացման գոտիները կոչվում են էուխրոմատին; քրոմոսոմների թերի թուլացում՝ հետերոքրոմատին: Քրոմատինը առավելագույնս խտանում է միտոտիկ բջիջների բաժանման ժամանակ, երբ այն հայտնաբերվում է խիտ քրոմոսոմների տեսքով։
Միջուկ. Սա 1-5 միկրոն չափի մեկ կամ մի քանի կլորացված մարմին է, որը ուժեղ բեկում է լույսը: Այն նաև կոչվում է նուկլեոլուս։ Միջուկը՝ միջուկի ամենախիտ կառուցվածքը, քրոմոսոմի ածանցյալն է։
Այժմ հայտնի է, որ միջուկը ցիտոպլազմայում ռիբոսոմային ՌՆԹ-ի և պոլիպեպտիդային շղթաների առաջացման վայրն է։
Միջուկն իր կառուցվածքով տարասեռ է. թեթև մանրադիտակով կարելի է տեսնել նրա մանրաթելային կազմակերպությունը։ Էլեկտրոնային մանրադիտակում առանձնանում են երկու հիմնական բաղադրիչ՝ հատիկավոր և մանրաթելային։ Ֆիբրիլային բաղադրիչը ռիբոսոմների պրեկուրսորների ռիբոնուկլեոպրոտեինային շղթաներն են, հատիկները՝ ռիբոսոմների հասունացման ենթամիավորներ։
Միջուկային ծրարը բաղկացած է արտաքին միջուկային թաղանթից և ծրարի ներքին թաղանթից՝ բաժանված պերինուկլեար տարածությամբ։ Միջուկային ծրարը պարունակում է միջուկային ծակոտիներ: Միջուկային թաղանթի թաղանթները մորֆոլոգիապես չեն տարբերվում մյուս ներբջջային թաղանթներից։
Ծակոտիները ունեն մոտ 80-90 նմ տրամագիծ։ Ծակոտի վրա դիֆրագմ կա: Այս բջջի ծակոտիների չափերը սովորաբար կայուն են: Ծակոտիների քանակը կախված է բջիջների նյութափոխանակության ակտիվությունից. որքան ավելի ինտենսիվ են սինթետիկ պրոցեսները բջիջներում, այնքան ավելի շատ ծակոտիներ են բջջի միջուկի միավորի մակերեսին:
Քրոմոսոմներ. Ե՛վ միջֆազային, և՛ միտոտիկ քրոմոսոմները բաղկացած են տարրական քրոմոսոմային մանրաթելերից՝ ԴՆԹ-ի մոլեկուլներից:
Միտոտիկ քրոմոսոմների մորֆոլոգիան լավագույնս ուսումնասիրվում է դրանց ամենամեծ խտացման պահին՝ մետաֆազում և անաֆազի սկզբում։ Այս վիճակում գտնվող քրոմոսոմները տարբեր երկարությամբ և բավականին հաստատուն հաստությամբ ձողաձև կառուցվածքներ են: Քրոմոսոմների մեծ մասում հեշտ է գտնել առաջնային կծկման գոտին (ցենտրոմեր), որը քրոմոսոմը բաժանում է երկու թեւերի։ Հավասար կամ գրեթե հավասար թեւեր ունեցող քրոմոսոմները կոչվում են մետակենտրոն, անհավասար երկարությամբ թեւեր՝ ենթամետասենտրիկ։ Շատ կարճ, գրեթե աննկատ երկրորդ թեւով ձողաձև քրոմոսոմները կոչվում են ակրոկենտրոն: Կինետոխորը գտնվում է առաջնային նեղացման շրջանում։ Բջջային լիսեռի միկրոխողովակները դուրս են գալիս այս գոտուց միտոզի ժամանակ: Որոշ քրոմոսոմներ ունեն նաև երկրորդական նեղացումներ, որոնք տեղակայված են քրոմոսոմի ծայրերից մեկի մոտ և բաժանում են փոքր տարածք՝ քրոմոսոմների արբանյակ: Այս վայրերում տեղայնացված է ռիբոսոմային ՌՆԹ-ի սինթեզի համար պատասխանատու ԴՆԹ-ն։
Քրոմոսոմների քանակի, չափերի և կառուցվածքային առանձնահատկությունների ամբողջությունը կոչվում է տվյալ տեսակի կարիոտիպ։ Տավարի կարիոտիպը 60 է, ձիերինը՝ 66, խոզերինը՝ 40, ոչխարներինը՝ 54, մարդունը՝ 46։
Բջջի գոյության ժամանակը որպես այդպիսին՝ բաժանումից բաժանում կամ բաժանումից մինչև մահ կոչվում է բջջային ցիկլ (նկ. 2):
Ամբողջ բջջային ցիկլը բաղկացած է 4 ժամանակաշրջանից՝ պատշաճ միտոզ, նախասինթետիկ, սինթետիկ և հետսինթետիկ ինտերֆազային շրջաններ: G1 ժամանակահատվածում բջիջների աճը սկսվում է բջջային սպիտակուցների կուտակման շնորհիվ, որը որոշվում է մեկ բջջի մեջ ՌՆԹ-ի քանակի ավելացմամբ: S - ժամանակահատվածում մեկ միջուկի վրա ԴՆԹ-ի քանակը կրկնապատկվում է և, համապատասխանաբար, քրոմոսոմների թիվը կրկնապատկվում է: Այստեղ ՌՆԹ-ի սինթեզի մակարդակը բարձրանում է ԴՆԹ-ի քանակի ավելացմանը համապատասխան՝ հասնելով առավելագույնին G2 շրջանում։ G2 ժամանակահատվածում սինթեզվում է սուրհանդակ ՌՆԹ, որն անհրաժեշտ է միտոզի անցման համար։ Այս պահին սինթեզված սպիտակուցների մեջ առանձնահատուկ տեղ են զբաղեցնում տուբուլինները՝ միտոտիկ spindle-ի սպիտակուցները։

Բրինձ. 2. Բջջի կյանքի ցիկլը.
M - միտոզ; G1 - նախասինթետիկ ժամանակաշրջան; S - սինթետիկ ժամանակաշրջան; G2 - հետսինթետիկ շրջան; 1 - հին բջիջ (2n4c); 2- երիտասարդ բջիջներ (2n2c)
Քրոմոսոմային հավաքածուի շարունակականությունն ապահովվում է բջիջների բաժանմամբ, որը կոչվում է միտոզ։ Այս գործընթացի ընթացքում տեղի է ունենում միջուկի ամբողջական վերակառուցում: Միտոզը բաղկացած է մի շարք փուլերից, որոնք հերթափոխվում են որոշակի հաջորդականությամբ՝ պրոֆազ, մետաֆազ, անաֆազ և տելոֆազ։ Միտոզի ընթացքում սոմատիկ բջջի միջուկը բաժանվում է այնպես, որ երկու դուստր բջիջներից յուրաքանչյուրը ստանում է քրոմոսոմների ճիշտ նույն շարքը, ինչ մայր բջիջը:
Բջիջների վերարտադրվելու ունակությունը կենդանի նյութի ամենակարեւոր հատկությունն է։ Այս ունակության շնորհիվ ապահովվում է բջիջների սերունդների շարունակական շարունակականություն, տեղի է ունենում բջջային կազմակերպվածության պահպանում կենդանիների էվոլյուցիայում, աճում և վերածնում։
Տարբեր պատճառներով (տրոհման spindle-ի խախտում, քրոմատիդների չբաժանում և այլն) բազմաթիվ օրգաններում և հյուսվածքներում հայտնաբերվում են մեծ միջուկներ կամ բազմամիջուկ բջիջներ ունեցող բջիջներ։ Սա սոմատիկ պոլիպլոիդիայի արդյունք է։ Այս երեւույթը կոչվում է էնդորարտադրություն: Պոլիպլոիդիան ավելի տարածված է անողնաշարավորների մոտ։ Նրանցից մի քանիսի մոտ տարածված է նաև պոլիթենիայի ֆենոմենը՝ ԴՆԹ-ի բազմաթիվ մոլեկուլներից քրոմոսոմի կառուցում։
Պոլիպլոիդ և պոլիտենային բջիջները չեն մտնում միտոզ և կարող են բաժանվել միայն ամիտոզով։ Իմաստը այս երեւույթըքանի որ և՛ պոլիպլոիդիան՝ քրոմոսոմների քանակի ավելացումը, և՛ պոլիթենիան՝ քրոմոսոմում ԴՆԹ մոլեկուլների քանակի ավելացումը, հանգեցնում են բջջի ֆունկցիոնալ ակտիվության զգալի աճի:
Բացի միտոզից, գիտությանը հայտնի է բաժանման ևս երկու տեսակ՝ ամիտոզ (և առանց, միտոզ՝ թելեր) կամ ուղղակի բաժանում և մեյոզ, որը քրոմոսոմների թիվը կիսով չափ կրճատելու գործընթաց է բջիջների երկու բաժանումներով՝ առաջին և երկրորդ։ մեյոզի բաժանում (meiosis - կրճատում): Մեյոզը բնորոշ է սեռական բջիջներին:
Գամետոգենեզ, վաղ սաղմի առաջացման փուլեր
1. Ողնաշարավորների սեռական բջիջների կառուցվածքը.
2. Սպերմատոգենեզ և ձվաբջիջ:
3. Վաղ սաղմի առաջացման փուլերը.
1. Սաղմնաբանություն՝ սաղմի զարգացման գիտություն։ Այն ուսումնասիրում է կենդանիների անհատական զարգացումը բեղմնավորման պահից (ձվի բեղմնավորումից) մինչև նրա ելքը կամ ծնունդը։ Սաղմնաբանությունը հաշվի է առնում սեռական բջիջների զարգացումն ու կառուցվածքը և սաղմի առաջացման հիմնական փուլերը՝ բեղմնավորում, տրոհում, գաստրուլյացիա, առանցքային օրգանների և օրգանոգենեզի, ժամանակավոր (ժամանակավոր) օրգանների զարգացում։
Ժամանակակից սաղմնաբանության ձեռքբերումները լայնորեն կիրառվում են անասնաբուծության, թռչնաբուծության և ձկնաբուծության մեջ; անասնաբուժության և բժշկության մեջ արհեստական բեղմնավորման և սերմնավորման, արագացված վերարտադրության և ընտրության տեխնոլոգիայի հետ կապված բազմաթիվ գործնական խնդիրների լուծման գործում. գյուղատնտեսական կենդանիների պտղաբերության բարձրացում, սաղմի փոխպատվաստման միջոցով կենդանիների բուծում, հղիության պաթոլոգիայի ուսումնասիրություն, անպտղության պատճառների ճանաչում և մանկաբարձական այլ խնդիրներ։
Կառուցվածքով սեռական բջիջները նման են սոմատիկ բջիջներին։ Դրանք նաև բաղկացած են միջուկից և ցիտոպլազմայից, որը կառուցված է օրգանելներից և ներդիրներից։
Հասուն գամետոցիտների տարբերակիչ հատկություններն են յուրացման և դիսիմիլացիոն գործընթացների ցածր մակարդակը, բաժանվելու անկարողությունը, քրոմոսոմների հապլոիդ (կես) քանակի միջուկներում պարունակությունը։
Բոլոր ողնաշարավորների արուների (սպերմատոզոիդների) սեռական բջիջները դրոշակաձև են (նկ. 3): Դրանք մեծ քանակությամբ ձևավորվում են ամորձիներում։ Մեկուսացված սերմնահեղուկի (սերմնաժայթքման) մի մասը պարունակում է տասնյակ միլիոնավոր և նույնիսկ միլիարդավոր սերմնաբջիջներ:
Գյուղատնտեսական կենդանիների սերմնահեղուկը շարժունակություն ունի։ Սերմնաբջիջների և՛ չափսերը, և՛ ձևը մեծապես տարբերվում են կենդանուց կենդանի: Դրանք բաղկացած են գլխից, պարանոցից և պոչից։ Սերմնաբջիջները տարասեռ են, քանի որ նրանց միջուկները պարունակում են տարբեր տեսակի սեռական քրոմոսոմներ: Սպերմատոզոիդների կեսն ունի X քրոմոսոմ, իսկ մյուս կեսը՝ Y քրոմոսոմ: Սեռական քրոմոսոմները կրում են գենետիկ տեղեկատվություն, որը որոշում է արական սեռի առանձնահատկությունները: Նրանք տարբերվում են այլ քրոմոսոմներից (ավտոսոմներից) իրենց հետերոքրոմատինի բարձր պարունակությամբ, չափերով և կառուցվածքով։
Սերմնաբջիջներն ունեն սննդանյութերի նվազագույն պաշար, որոնք շատ արագ սպառվում են, երբ բջիջը շարժվում է: Եթե սերմնահեղուկը չի միաձուլվում ձվի հետ, ապա այն սովորաբար մահանում է կանանց սեռական տրակտում 24-36 ժամ հետո։
Դուք կարող եք երկարացնել սերմնահեղուկի կյանքը սառեցնելով։ Քինինը, ալկոհոլը, նիկոտինը և այլ դեղամիջոցներ վնասակար ազդեցություն ունեն սերմի վրա։
Ձվի կառուցվածքը. Ձվաբջիջը շատ ավելի մեծ է, քան սերմնահեղուկը: Ձվաբջիջների տրամագիծը տատանվում է 100 մկմ-ից մինչև մի քանի մմ: Ողնաշարավորների ձվերը օվալաձև են, անշարժ, կազմված են միջուկից և ցիտոպլազմայից (նկ. 4): Միջուկը պարունակում է քրոմոսոմների հապլոիդ հավաքածու։ Կաթնասունների ձվերը դասակարգվում են որպես հոմոգամետիկ, քանի որ դրանց միջուկը պարունակում է միայն X քրոմոսոմը: Ցիտոպլազմը պարունակում է ազատ ռիբոսոմներ, էնդոպլազմիկ ցանց, Գոլջիի բարդույթ, միտոքոնդրիա, դեղնուց և այլ բաղադրիչներ։ Ձվաբջիջները բևեռային են: Դրանցում առանձնանում են երկու բևեռներ՝ գագաթային և բազալ։ Ձվի ցիտոպլազմայի ծայրամասային շերտը կոչվում է կեղևային շերտ (կեղև - կեղև): Այն ամբողջովին զուրկ է դեղնուցից, պարունակում է բազմաթիվ միտոքոնդրիաներ։
Ձվերը ծածկված են թաղանթներով։ Տարբերում են առաջնային, երկրորդային և երրորդային թաղանթներ։ Առաջնային շերտը պլազմալեման է: Երկրորդային թաղանթը (թափանցիկ կամ փայլուն) ձվարանների ֆոլիկուլյար բջիջների ածանցյալն է։ Թռչունների ձվաբջիջում ձևավորվում են երրորդային թաղանթներ՝ ձվի սպիտակուցը, կեղևը և թաղանթները: Դեղնուցի քանակով ձուն առանձնանում է փոքր քանակությամբ՝ օլիգոլեցիտալ (օլիգոս՝ քիչ, լեցիտ՝ դեղնուց), միջին քանակով՝ մեսոլեցիտալ (մեսոս՝ միջին) և մեծ քանակությամբ՝ պոլիլեցիտալ (պոլիս՝ շատ)։
Ըստ ցիտոպլազմայում դեղնուցի տեղակայման՝ ձվերը առանձնանում են դեղնուցի միատեսակ բաշխմամբ՝ իզոլեցիտալ կամ հոմոլեցիտալ, իսկ դեղնուցի տեղայնացումով մեկ բևեռում՝ տելոլեցիտալ (տելոս՝ ծայր, ծայր)։ Օլիգոլեցիտալ և իզոլեցիտալ ձվերը՝ նշտարակի և կաթնասունների մեջ, մեզոլեցիտալ և տելոլեցիտալ՝ երկկենցաղներում, որոշ ձկներ, պոլիլեցիտալ և տելոլեցիտալ՝ շատ ձկների, սողունների, թռչունների մոտ։
2. Սեռական բջիջների նախնիները առաջնային սեռական բջիջներն են՝ գամետոբլաստները (գոնոբլաստները): Դրանք հայտնաբերվում են արյունատար անոթների մոտ գտնվող դեղնուցի պարկի պատում։ Գոնոբլաստները ինտենսիվորեն բաժանվում են միտոզով և արյան հոսքով կամ արյունատար անոթների ընթացքով գաղթում են դեպի սեռական գեղձերի սկզբնական հատվածները, որտեղ դրանք շրջապատված են օժանդակ (ֆոլիկուլյար) բջիջներով։ Վերջիններս կատարում են տրոֆիկ ֆունկցիա։ Այնուհետև, կապված կենդանու սեռի զարգացման հետ, սեռական բջիջները ձեռք են բերում սերմնաբջիջներին և ձվաբջիջներին բնորոշ հատկություններ։
Սերմնահեղուկի զարգացումը (սպերմատոգենեզ) տեղի է ունենում սեռական հասուն կենդանու ամորձիներում։ Սպերմատոգենեզում կա 4 շրջան՝ վերարտադրություն, աճ, հասունացում և ձևավորում։
բուծման շրջանը. Բջիջները կոչվում են սպերմատոգոնիա: Նրանք փոքր են և ունեն քրոմոսոմների դիպլոիդ քանակ։ Բջիջները արագորեն բաժանվում են միտոզով: Բաժանվող բջիջները ցողունային բջիջներ են և լրացնում են սպերմատոգոնիայի պաշարը։
Աճի ժամանակաշրջան. Բջիջները կոչվում են առաջնային սպերմատոցիտներ: Նրանք ունեն քրոմոսոմների դիպլոիդ քանակ։ Բջջի չափը մեծանում է, և միջուկում ժառանգական նյութի վերաբաշխման մեջ տեղի են ունենում բարդ փոփոխություններ, որոնց հետ կապված առանձնանում են չորս փուլեր՝ լեպտոտենային, զիգոտենային, պախիտային, դիպլոտենային։
Հասունացման շրջան. Սա քրոմոսոմների կես թվով սերմնաբջիջների զարգացումն է:
Հասունացման գործընթացում յուրաքանչյուր առաջնային սպերմատոցիտից առաջանում են 4 սպերմատիդներ՝ մեկ թվով քրոմոսոմներով։ Ունեն լավ զարգացած միտոքոնդրիաներ, Գոլջիի բարդույթ, ցենտրոսոմ՝ տեղակայված միջուկի մոտ։ Մյուս օրգանելները, ինչպես նաև ներդիրները, գրեթե բացակայում են։ Սպերմատիդները չեն կարողանում բաժանվել։
Կազմավորման շրջանը. Սպերմատիդը ձեռք է բերում սերմնահեղուկին բնորոշ մորֆոլոգիական հատկություններ։ Գոլջիի կոմպլեքսը վերածվում է ակրոսոմի, որը գլխարկի տեսքով պարփակում է սերմնաբջիջի միջուկը։ Ակրոսոմը հարուստ է հիալուրոնիդազ ֆերմենտով։ Ցենտրոսոմը միջուկից շարժվում է դեպի հակառակ բևեռը, որում տարբերվում են պրոքսիմալ և դիստալ ցենտրիոլները։ Պրոքսիմալ ցենտրիոլը մնում է սերմի պարանոցում, մինչդեռ հեռավոր ցենտրիոլը գնում է պոչը կառուցելու:
Ձվի, ձվաբջիջների զարգացումը բարդ և շատ երկար գործընթաց է: Այն սկսվում է էմբրիոգենեզի ժամանակաշրջանում և ավարտվում սեռական հասուն կնոջ վերարտադրողական համակարգի օրգաններում։ Օոգենեզը բաղկացած է երեք շրջանից՝ վերարտադրություն, աճ, հասունացում:
Վերարտադրողական շրջանը տեղի է ունենում ներարգանդային զարգացման շրջանում և ավարտվում ծնվելուց հետո առաջին ամիսների ընթացքում։ Բջիջները կոչվում են օվոգոնիա և ունեն քրոմոսոմների դիպլոիդ քանակ։
Աճման ընթացքում բջիջները կոչվում են առաջնային ձվաբջիջներ: Միջուկների փոփոխությունները նման են առաջնային սպերմատոցիտների: Այնուհետև ձվաբջիջում սկսվում է դեղնուցի ինտենսիվ սինթեզը և կուտակումը` նախավիտելլոգենեզի փուլը և վիտելոգենեզի փուլը։ Ձվաբջիջի երկրորդական թաղանթը բաղկացած է ֆոլիկուլյար բջիջների մեկ շերտից: Նախավիտելոգենեզը սովորաբար տևում է մինչև իգական սեռի հասունացումը: Հասունացման շրջանը բաղկացած է արագ հաջորդական հասունացման բաժանումներից, որոնց ընթացքում դիպլոիդ բջիջը դառնում է հապլոիդ։ Այս գործընթացը սովորաբար տեղի է ունենում ձվաբջջի մեջ՝ օվուլյացիայից հետո:
Հասունացման առաջին բաժանումն ավարտվում է երկու անհավասար կառուցվածքների՝ երկրորդական ձվաբջիջի և առաջին ուղղորդված կամ ռեդուկցիոն մարմնի ձևավորմամբ։ Երկրորդ բաժանման ժամանակ ձևավորվում է նաև մեկ հասուն ձու և երկրորդ ուղղորդված մարմին։ Առաջին մարմինը նույնպես բաժանված է. Հետևաբար, հասունացման ընթացքում մեկ առաջնային ձվաբջիջից առաջանում է միայն մեկ հասուն ձվաբջիջ, և վերջինիս երեք ուղղորդված մարմինները շուտով մահանում են։
Բոլոր ձվերը գենետիկորեն միատարր են, քանի որ ունեն միայն X քրոմոսոմ:
3. Բեղմնավորում - սեռական գամետների միաձուլում և նոր միաբջիջ օրգանիզմի (զիգոտի) ձևավորում։ Այն տարբերվում է հասուն ձվաբջիջից ԴՆԹ-ի կրկնակի զանգվածով՝ քրոմոսոմների դիպլոիդ քանակով։ Կաթնասունների մոտ բեղմնավորումը ներքին է, այն տեղի է ունենում ձվաբջիջում իր պասիվ շարժումով դեպի արգանդ։ Սպերմատոզոիդների տեղաշարժը կնոջ սեռական տրակտում իրականացվում է այս բջջի շարժման ապարատի ֆունկցիայի (քիմոտաքսիս և ռեոտաքսիս), արգանդի պատի պերիստալտիկ կծկումների և ձվաբջիջի ներքին մակերեսը ծածկող թարթիչների շարժման շնորհիվ: . Երբ սեռական բջիջները մոտենում են միմյանց, սերմնաբջիջների գլխի ակրոսոմի ֆերմենտները ոչնչացնում են ֆոլիկուլյար բջիջների շերտը՝ ձվի երկրորդական թաղանթը։ Այն պահին, երբ սերմնահեղուկը դիպչում է ձվի պլազմոլեմային, դրա մակերեսին առաջանում է ցիտոպլազմայի ելուստ՝ բեղմնավորման տուբերկուլյոզ։ Գլուխն ու պարանոցը թափանցում են ձվաբջիջի մեջ։ Կաթնասունների մոտ միայն մեկ սերմնահեղուկ է մասնակցում բեղմնավորմանը, հետևաբար, գործընթացը կոչվում է մոնոսպերմիա՝ XY՝ արու, XX՝ էգ:
Թռչուններն ու սողունները ունեն պոլիսպերմիա։ Թռչունների մոտ բոլոր սերմնահեղուկներն ունեն Z-քրոմոսոմ, իսկ ձվերը՝ Z կամ W-քրոմոսոմ:
Սերմնահեղուկի ձվաբջիջ ներթափանցելուց հետո վերջինիս շուրջ գոյանում է բեղմնավորման թաղանթ՝ կանխելով այլ սերմնաբջիջների ներթափանցումը ձվաբջիջ, սեռական բջիջների միջուկները կոչվում են՝ արական պրոնուկլուս, իգական պրոնուկլուս։ Նրանց միացման գործընթացը կոչվում է սինկարիոն։ Սերմնահեղուկի կողմից ներմուծված ցենտրիոլը բաժանվում և շեղվում է՝ ձևավորելով ախրոմատինային ողորկ։ Ջախջախումը սկսվում է: Ճեղքումը միաբջիջ զիգոտի զարգացման հետագա գործընթացն է, որի ընթացքում ձևավորվում է բազմաբջիջ բլաստուլա, որը բաղկացած է պատից՝ բլաստոդերմից և խոռոչից՝ բլաստոկոելից։ Զիգոտի միտոտիկ բաժանման գործընթացում ձևավորվում են նոր բջիջներ՝ բլաստոմերներ։
Քորդատներում մասնատման բնույթը տարբեր է և մեծապես պայմանավորված է ձվի տեսակով։ Ճեղքվածքը կարող է լինել ամբողջական (հոլոբլաստիկ) կամ մասնակի (մերոբլաստիկ): Առաջին տիպում մասնակցում է զիգոտի ամբողջ նյութը, երկրորդում՝ նրա միայն այն գոտին, որը զուրկ է դեղնուցից։
Ամբողջական ջախջախումը դասակարգվում է միատարր և անհավասար: Առաջինը բնորոշ է օլիգոիզոլեցիտալ ձվերին (նշտարակ, կլոր որդ և այլն)։ Բեղմնավորված ձվի մեջ առանձնանում են երկու բևեռներ՝ վերինը՝ կենդանական և ստորինը՝ վեգետատիվ։ Բեղմնավորումից հետո դեղնուցը տեղափոխվում է վեգետատիվ բևեռ։
Ճեղքվածքն ավարտվում է բլաստուլայի առաջացմամբ, որի ձևը հեղուկով լցված գնդակ է հիշեցնում։ Գնդիկի պատը ձևավորվում է բլաստոդերմային բջիջներով: Այսպիսով, ամբողջական միատեսակ ճեղքումով, ամբողջ զիգոտի նյութը մասնակցում է ճեղքմանը, և յուրաքանչյուր բաժանումից հետո բջիջների թիվը կրկնապատկվում է։
Լրիվ անհավասար ճեղքվածքը բնորոշ է մեզոլեցիտալ (դեղնուցի միջին քանակություն) և տելոլեցիտալ ձվաբջիջներին։ Սրանք երկկենցաղներ են: Նրանց տեսակը blastula է coeloblastula.
Մասնակի կամ մերոբլաստիկ (դիսկոիդային) ճեղքումը տարածված է ձկների, թռչունների մոտ և բնորոշ է պոլիլեցիտալ և տելոլեցիտալ ձվերին (բլաստուլայի տեսակը կոչվում է դիսկոբլաստուլա)։
Գաստրուլյացիա. Բլաստուլայի հետագա զարգացմամբ, բջիջների բաժանման, աճի, տարբերակման և դրանց շարժումների գործընթացում սկզբում ձևավորվում է երկշերտ սաղմ։ Նրա շերտերն են էկտոդերմը, էնդոդերմը և մեզոդերմը։
Գաստրուլյացիայի տեսակները. 1) ինվագինացիա, 2) էպիբոլիա (աղտոտում), 3) ներգաղթ (տեղաբաշխում), 4) շերտավորում (շերտավորում):
Էջանշեք առանցքային օրգանները: Այս սաղմնային շերտերից ձևավորվում են առանցքային օրգաններ՝ սաղմը նյարդային համակարգ(նյարդային խողովակ), նոտոկորդ և աղիքային խողովակ:
Մեզոդերմի զարգացման գործընթացում բոլոր ողնաշարավորները ձևավորում են նոտոկորդ, հատվածավորված մեզոդերմա կամ սոմիտներ (ողնաշարի հատվածներ) և չհատված մեզոդերմա կամ սպլանխնոտ։ Վերջինս բաղկացած է երկու թիթեղից՝ արտաքին՝ պարիետալ և ներքին՝ վիսցերալ։ Այս թիթեղների միջև ընկած տարածությունը կոչվում է մարմնի երկրորդական խոռոչ:
Սոմիտներում առանձնանում են երեք ռուդիմենտներ՝ դերմատոմ, միոտոմ, սկլերոտոմ։ Նեֆրոգոնադոտոմ.
Սաղմնային շերտերի տարբերակմամբ առաջանում է սաղմնային հյուսվածք՝ մեզենխիմ։ Այն զարգանում է հիմնականում մեզոդերմայից և էկտոդերմայից գաղթած բջիջներից։ Մեզենխիմը կապի հյուսվածքի, հարթ մկանների, արյան անոթների և կենդանիների մարմնի այլ հյուսվածքների զարգացման աղբյուր է։ Քորդատների տարբեր ներկայացուցիչների մոտ տրոհման պրոցեսները շատ յուրահատուկ են և կախված են ձվի պրոմորֆոլոգիայից, հատկապես դեղնուցի քանակից և բաշխումից։ Գաստրուլյացիայի գործընթացները նույնպես մեծապես տարբերվում են Chordata-ում:
Այսպիսով, նշտարում գաստրուլյացիան սովորաբար ինվագինացիա է, այն սկսվում է ենթադրյալ էնդոդերմի ինվագինացիայից: Էնդոդերմայից հետո նոտոկորդային նյութը ներխուժում է բլաստոկոել, և մեզոդերմը սուզվում է բլաստոպորի կողային և փորային շուրթերով: Բլաստոպորի առջևի (կամ մեջքի) շրթունքը բաղկացած է ապագա նյարդային համակարգի նյութից, իսկ ներսից՝ ապագա նոտոկորդի բջիջներից։ Հենց էնդոդերմալ շերտը շփվում է էկտոդերմալ շերտի ներքին կողմի հետ, սկսվում են գործընթացներ, որոնք հանգեցնում են առանցքային օրգանների պրիմորդիայի առաջացմանը։
Ոսկրային ձկների մոտ գաստրուլյացիայի պրոցեսը սկսվում է այն ժամանակ, երբ բազմաշերտ բլաստոդսկը ծածկում է ձվի դեղնուցի միայն մի փոքր մասը և ավարտվում է ամբողջ «դեղնուցի գնդիկի» ամբողջական աղտոտմամբ։ Սա նշանակում է, որ գաստրուլյացիան ներառում է նաև բլաստոդիսկի աճը։
Բլաստոդսկի առաջի և կողային եզրերի երկայնքով բոլոր երեք սաղմնային շերտերի բջջային նյութը սկսում է աճել դեղնուցի վրա: Այսպիսով, ձևավորվում է այսպես կոչված դեղնուցի պարկը։
Դեղնուցային պարկը, որպես սաղմի մի մաս, կատարում է մի շարք գործառույթներ.
1) դա տրոֆիկ ֆունկցիա ունեցող օրգան է, քանի որ տարբերակիչ էնդոդերմալ շերտը արտադրում է ֆերմենտներ, որոնք օգնում են քայքայել դեղնուցի նյութերը, իսկ տարբերակիչ մեզոդերմալ շերտում ձևավորվում են արյունատար անոթներ, որոնք կապված են սաղմի անոթային համակարգի հետ։ ինքն իրեն։
2) դեղնուցի պարկը շնչառական օրգան է. Սաղմի գազի փոխանակումը արտաքին միջավայրի հետ տեղի է ունենում պարկի անոթների և էկտոդերմալ էպիթելի պատերի միջոցով։
3) «արյան մեզենխիմը» արյունաստեղծության բջջային հիմքն է։ Դեղնապարկը սաղմի առաջին արյունաստեղծ օրգանն է։
Գորտեր, տրիտոններ և ծովային ոզնիներքսաներորդ դարի փորձարարական սաղմնային հետազոտության հիմնական օբյեկտներն են։
Երկկենցաղների մոտ ներխուժումը չի կարող տեղի ունենալ այնպես, ինչպես նշտարում, քանի որ ձվի վեգետատիվ կիսագունդը շատ ծանրաբեռնված է դեղնուցով։
Գորտերի մոտ սկզբնական գաստրուլյացիայի առաջին նկատելի նշանը բլաստոպորի առաջացումն է, այսինքն՝ գորշ կիսալուսնի մեջտեղում անցք կամ բացվածք:
Առանձնահատուկ ուշադրության է արժանի նյարդային համակարգի և մաշկի էպիդերմիսի բջջային նյութի վարքը։ Ի վերջո, նյարդային համակարգի ապագա էպիդերմիսը և նյութը ծածկում են սաղմի ամբողջ մակերեսը: Մաշկի ենթադրյալ էպիդերմիսը շարժվում և նոսրանում է բոլոր ուղղություններով: Ենթադրյալ նյարդային համակարգի բջիջների ամբողջությունը շարժվում է գրեթե բացառապես միջօրեական ուղղություններով։ Ապագա նյարդային համակարգի բջիջների շերտը լայնակի ուղղությամբ կրճատվում է, նյարդային համակարգի ենթադրյալ շրջանը երկարացվում է կենդանի-վեգետատիվ ուղղությամբ։
Եկեք ամփոփենք այն, ինչ գիտենք սաղմնային շերտերից յուրաքանչյուրի ճակատագրի մասին:
Էկտոդերմի ածանցյալներ. Արտաքին շերտը կազմող բջիջներից, բազմապատկելով և տարբերվող, ձևավորվում են՝ արտաքին էպիթելի, մաշկային գեղձեր, ատամների մակերեսային շերտ, եղջյուրավոր թեփուկներ և այլն։ Ի դեպ, գրեթե միշտ յուրաքանչյուր օրգան զարգանում է երկու բջջային տարրերից։ կամ նույնիսկ բոլոր երեք սաղմնային շերտերը: Օրինակ, կաթնասունների մաշկը զարգանում է էկտոդերմայից և մեզոդերմայից:
Առաջնային էկտոդերմայի ընդարձակ հատվածը «սուզվում է» դեպի ներս՝ արտաքին էպիթելի տակ և առաջացնում է ամբողջ նյարդային համակարգը։
Էնդոդերմի ածանցյալներ. Ներքին սաղմնային շերտը զարգանում է միջին աղիքի էպիթելի և նրա մարսողական գեղձերի մեջ: Շնչառական համակարգի էպիթելը զարգանում է առաջի աղիքից։ Բայց դրա ծագման մեջ ներգրավված է այսպես կոչված նախակորդալ ափսեի բջջային նյութը:
մեզոդերմի ածանցյալներ. Դրանից զարգանում են բոլոր մկանային հյուսվածքները, շարակցական, աճառային, ոսկրային հյուսվածքների բոլոր տեսակները, արտազատման օրգանների ուղիները, մարմնի խոռոչի որովայնի խոռոչը, շրջանառու համակարգը, ձվարանների և ամորձիների հյուսվածքների մի մասը։
Կենդանիների մեծ մասում միջին շերտը հայտնվում է ոչ միայն բջիջների հավաքածուի տեսքով, որոնք կազմում են կոմպակտ էպիթելային շերտ, այսինքն՝ համապատասխան մեզոդերմա, այլև ցրված, ամեոբանման բջիջների ազատ համալիրի տեսքով: Մեզոդերմի այս հատվածը կոչվում է մեզենխիմ: Իրականում մեզոդերմը և մեզենխիմը տարբերվում են միմյանցից իրենց ծագմամբ, նրանց միջև ուղղակի կապ չկա, հոմոլոգ չեն։ Մեզենխիմը հիմնականում էկտոդերմալ ծագում ունի, մինչդեռ մեզոդերմը առաջանում է էնդոդերմայից։ Ողնաշարավորների մոտ, սակայն, մեզենխիմը ընդհանուր ծագում ունի մեսոդերմի մնացած մասերի հետ։
Բոլոր կենդանիների մոտ, որոնք հակված են կոելոմի (երկրորդային մարմնի խոռոչ) ունենալուն, մեզոդերմից առաջանում են սնամեջ պարկեր: Կոելոմիկ պարկերը ձևավորվում են սիմետրիկորեն աղիքի կողքերին: Յուրաքանչյուր կոելոմիկ պարկի պատը, որը ուղղված է դեպի աղիքներ, կոչվում է splanchnopleura: Սաղմի էկտոդերման դեմ ուղղված պատը կոչվում է սոմատոպլեուրա։
Այսպիսով, սաղմի զարգացման ընթացքում ձևավորվում են տարբեր խոռոչներ, որոնք ունեն կարևոր մորֆոգենետիկ նշանակություն։ Սկզբում հայտնվում է Բաերի խոռոչը՝ վերածվելով մարմնի առաջնային խոռոչի՝ բլաստոկոելի, ապա առաջանում է գաստրոկոելը (կամ ստամոքսի խոռոչը) և վերջապես, շատ կենդանիների մոտ՝ ամբողջը։ Գաստրոկոելի և կոելոմի ձևավորմամբ բլաստոկոելը ավելի ու ավելի է նվազում, այնպես որ նախկին առաջնային մարմնի խոռոչից մնում են միայն աղիքի պատերի և կոելոմի միջև եղած բացերը։ Այս բացերը վերածվում են շրջանառության համակարգի խոռոչների։ Գաստրոկոելն ի վերջո վերածվում է միջին աղիքի խոռոչի:
Կաթնասունների և թռչունների սաղմնավորման առանձնահատկությունները
1. Արտսաղմնային օրգաններ.
2. Կաթնասունների պլասենտա.
3. Որոճողների, խոզերի և թռչունների նախածննդյան օնտոգենեզի փուլերը.
1. Սողունների և թռչունների սաղմերում ձևավորվում է նաև դեղնուցի պարկ։ Սրանում ներգրավված են բոլոր մանրէային շերտերը: Հավի սաղմի զարգացման 2-րդ և 3-րդ օրերի ընթացքում opaca-ի ներքին հատվածում զարգանում է արյունատար անոթների ցանց։ Նրանց տեսքը անքակտելիորեն կապված է սաղմնային արյունաստեղծության առաջացման հետ: Այսպիսով, թռչնի սաղմերի դեղնուցային պարկի գործառույթներից մեկը սաղմնային արյունաստեղծումն է։ Սաղմի մեջ միայն այնուհետև ձևավորվում են արյունաստեղծ օրգաններ՝ լյարդ, փայծաղ, ոսկրածուծ։
Պտղի սիրտը սկսում է գործել (կծկվել) երկրորդ օրվա վերջում, այդ պահից արյան հոսք կա։
Թռչունների սաղմերում, բացի դեղնուցի պարկից, ձևավորվում են ևս երեք ժամանակավոր օրգաններ, որոնք սովորաբար կոչվում են սաղմնային թաղանթներ՝ ամնիոն, սերոզա և ալանտոիս։ Այս օրգանները կարելի է համարել զարգացած սաղմերի էվոլյուցիոն ադապտացիայի գործընթացում։
Ամնիոնը և սերոզան առաջանում են ամենասերտ հարաբերություններում: Ամնիոն լայնակի ծալքի տեսքով, աճող, թեքվում է սաղմի գլխի առջևի ծայրի վրա և ծածկում է այն գլխարկի պես։ Ապագայում ամնիոտիկ ծալքերի կողային հատվածները աճում են հենց սաղմի երկու կողմերում և միասին աճում: Ամնիոտիկ ծալքերը կազմված են էկտոդերմայից և պարիետալ մեզոդերմայից։
Ամնիոտիկ խոռոչի պատի հետ մեկտեղ զարգանում է մեկ այլ կարևոր ժամանակավոր ձևավորում՝ շիճուկը կամ շիճուկային թաղանթը: Այն բաղկացած է էկտոդերմալ տերևից, որը «նայում է» սաղմին, և մեզոդերմայից՝ «նայող» դեպի դուրս։ Արտաքին կեղևը աճում է կեղևի տակ գտնվող ամբողջ մակերեսով: Սա սերոզա է:
Ամնիոնը և սերոզան, իհարկե, «պատյաններ» են, քանի որ դրանք իսկապես ծածկում և միավորում են հենց սաղմը արտաքին միջավայրից։ Այնուամենայնիվ, դրանք օրգաններ են, սաղմի մասեր, որոնք ունեն շատ կարևոր գործառույթներ։ Ամնիոտիկ հեղուկը ջրային միջավայր է ստեղծում կենդանիների սաղմերի համար, որոնք էվոլյուցիայի ընթացքում դարձել են ցամաքային: Այն պաշտպանում է զարգացող սաղմը չորանալուց, ցնցումից, ձվի կեղևին կպչելուց։ Հետաքրքիր է նշել, որ ամնիոտիկ հեղուկի դերը կաթնասունների մեջ նշել է Լեոնարդո դա Վինչին։
Շիճուկային թաղանթը մասնակցում է սպիտակուցային մեմբրանի մնացորդների շնչառությանը և ռեզորբմանը (խորիոնի կողմից արտազատվող ֆերմենտների ազդեցության տակ):
Զարգանում է մեկ այլ ժամանակավոր օրգան՝ ալանտոիսը, որն առաջին հերթին կատարում է սաղմնային միզապարկի ֆունկցիան։ Այն հայտնվում է որպես հետին աղիքային էնդոդերմի որովայնային ելք: Հավի սաղմի մեջ այս ելուստը հայտնվում է արդեն զարգացման 3-րդ օրը։ Թռչունների սաղմնային զարգացման միջնամասում ալանտոիսն աճում է քորիոնի տակ սաղմի ամբողջ մակերեսով դեղնուցի պարկով:
Թռչունների (և սողունների) սաղմնային զարգացման ամենավերջում սաղմի ժամանակավոր օրգանները աստիճանաբար դադարեցնում են իրենց գործառույթները, դրանք կրճատվում են, սաղմը սկսում է շնչել օդը ձվի ներսում (օդային խցիկում), ճեղքում է կեղևը, ազատվում է ձվի թաղանթներից և հայտնվում արտաքին միջավայրում։
Կաթնասունների արտասաղմնային օրգաններն են՝ դեղնուցի պարկը, ամնիոնը, ալանտոիզը, քորիոնը և պլասենտան (նկ. 5):
2. Կաթնասունների մոտ սաղմի կապը մոր մարմնի հետ ապահովվում է հատուկ օրգանի՝ պլասենցայի (մանկական տեղ) գոյացմամբ։ Նրա զարգացման աղբյուրը ալանտո-քորիոնն է։ Պլասենտաները ըստ կառուցվածքի բաժանվում են մի քանի տեսակների. Դասակարգումը հիմնված է երկու սկզբունքի վրա՝ ա) խորիոնային վիլլիների բաշխման բնույթը և 2) արգանդի լորձաթաղանթի հետ կապվածության ձևը (նկ. 6):
Ձևը առանձնացնում է պլասենցայի մի քանի տեսակներ.
1) ցրված պլասենտա (էպիթելիոխորիալ) - նրա երկրորդական պապիլները զարգանում են խորիոնի ամբողջ մակերեսով: Քորիոնիկ վիլլիները ներթափանցում են արգանդի պատի գեղձերը՝ առանց արգանդի հյուսվածքի ոչնչացման։ Սաղմի սնուցումն իրականացվում է արգանդի գեղձերի միջոցով, որոնք արտազատում են արքայական ժելե, որը ներծծվում է խորիոնային վիլլի արյունատար անոթների մեջ։ Ծննդաբերության ժամանակ խորիոնային վիլլիները դուրս են մղվում արգանդի գեղձերից՝ առանց հյուսվածքների քայքայման։ Նման պլասենտան բնորոշ է խոզերի, ձիերի, ուղտերի, մարսուների, կետաձկանների, գետաձիու համար։

Բրինձ. 5. Կաթնասունների դեղնուցի պարկի և սաղմնային թաղանթների զարգացման սխեման (վեց հաջորդական փուլ).
Ա - պտղի միզապարկի խոռոչի աղտոտման գործընթացը էնդոդերմայով (1) և մեզոդերմայով (2); B - փակ էնդոդերմալ վեզիկուլի ձևավորում (4); B - ամնիոտիկ ծալքի (5) և աղիքային ակոսի ձևավորման սկիզբը (6); G - սաղմի մարմնի մեկուսացում (7); դեղնուցի պարկ (8); D - ամնիոտիկ ծալքերի փակում (9); ալլանտոիսի զարգացման ձևավորման սկիզբը (10); E - փակ ամնիոտիկ խոռոչ (11); զարգացած allantois (12); chorionic villi (13); Մեզոդերմի պարիետալ թերթիկ (14); Մեզոդերմի վիսցերալ թերթիկ (15); էկտոդերմա (3).
2) Կոթիլեդոնային պլասենտա (դեսմոխորիոնիկ) - խորիոնային վիլլիները գտնվում են թփուտներում՝ կոթիլեդոններում։ Դրանք կապվում են արգանդի պատի խտացումների հետ, որոնք կոչվում են կարունկուլներ։ Կոթիլեդոն-կարունկային համալիրը կոչվում է պլասենտա: Նման պլասենտան բնորոշ է որոճողների համար։
3) Գոտու պլասենտա (էնդոթելիոխորիոնիկ) - լայն գոտու տեսքով վիլիները շրջապատում են պտղի միզապարկը և գտնվում են արգանդի պատի շարակցական հյուսվածքի շերտում՝ արյան անոթների պատի էնդոթելիային շերտի հետ շփման մեջ։
4) Դիսկոիդային պլասենտա (հեմոխորիալ) - խորիոնային վիլլիի և արգանդի պատի շփման գոտին ունի սկավառակի ձև: Քորիոնիկ վիլլիները սուզվում են արյունով լցված բացվածքների մեջ, որոնք ընկած են արգանդի պատի շարակցական հյուսվածքի շերտում: Այս պլասենտան հանդիպում է պրիմատների մոտ:
3. Անասնաբուծության աշխատողներն իրենց գործնական գործունեությամբ զբաղվում են կենդանիների բուծմամբ և բուծմամբ: Սրանք բարդ կենսաբանական գործընթացներ են, և դրանք գիտակցաբար կառավարելու կամ դրանք բարելավելու ուղիներ փնտրելու համար կենդանաբանական այգու ինժեներն ու անասնաբույժը պետք է իմանան կենդանիների զարգացման հիմնական օրինաչափությունները իրենց անհատական կյանքի ընթացքում: Մենք արդեն գիտենք, որ փոփոխությունների շղթան, որի միջով անցնում է օրգանիզմը սկզբից մինչև բնական մահ, կոչվում է օնտոգեն: Այն կազմված է որակապես տարբեր ժամանակաշրջաններից։ Այնուամենայնիվ, օնտոգենեզի պարբերականացումը դեռ բավականաչափ զարգացած չէ: Որոշ գիտնականներ կարծում են, որ օրգանիզմի օնտոգենետիկ զարգացումը սկսվում է սեռական բջիջների, մյուսները՝ զիգոտի ձևավորմամբ։

Բրինձ. 6. Պլասենցայի հյուսվածքաբանական կառուցվածքի տեսակները.
A - epitheliochorial; B - desmochorial; C - էնդոթելիոխորիալ, G - հեմոխորիալ; I - բողբոջային մաս; II - մայրական մաս; 1 - էպիթելիա. 2 - շարակցական հյուսվածք և 3 - քորիոնիկ վիլուսի արյունատար անոթի էնդոթելիում; 4 - էպիթելիա; 5 - շարակցական հյուսվածք և 6 - արյան անոթներ և արգանդի լորձաթաղանթի բացվածքներ:
Զիգոտի հայտնվելուց հետո գյուղատնտեսական կենդանիների հետագա օնտոգենեզը բաժանվում է ներարգանդային և հետծննդյան զարգացման։
Գյուղատնտեսական կենդանիների ներարգանդային զարգացման ենթաշրջանների տեւողությունը, օրեր (ըստ Գ.Ա. Շմիդտի).
Կենդանիների սաղմնավորման մեջ, իրենց փոխհարաբերությունների շնորհիվ, կան մի քանի սկզբունքորեն նման հատկանիշներ. հյուսվածքներ և օրգաններ.
Ընդհանուր հյուսվածաբանություն. էպիթելային հյուսվածքներ
1. Հյուսվածքների զարգացում.
2. Էպիթելային հյուսվածքների դասակարգում.
3. Խցուկներ և դրանց դասակարգման չափանիշներ.
1. Կենդանիների մարմինը կառուցված է բջիջներից և ոչ բջջային կառուցվածքներից, որոնք մասնագիտացած են որոշակի գործառույթներ կատարելու մեջ: Բջիջների պոպուլյացիաները՝ տարբեր ֆունկցիաներով, տարբերվում են ներբջջային սպիտակուցների սինթեզի կառուցվածքով և յուրահատկությամբ։
Զարգացման գործընթացում ի սկզբանե միատարր բջիջները ձեռք են բերել նյութափոխանակության, կառուցվածքի և ֆունկցիայի տարբերություններ։ Այս գործընթացը կոչվում է տարբերակում: Այս դեպքում գիտակցվում է գենետիկ տեղեկատվությունը, որը բխում է բջջի միջուկի ԴՆԹ-ից, որն արտահայտվում է կոնկրետ պայմաններում։ Բջիջների հարմարեցումը այս պայմաններին կոչվում է հարմարվողականություն:
Տարբերակումը և հարմարեցումը որոշում են բջիջների և դրանց պոպուլյացիաների միջև որակապես նոր փոխհարաբերությունների և հարաբերությունների զարգացումը: Միևնույն ժամանակ մեծապես մեծանում է օրգանիզմի ամբողջականության, այսինքն՝ ինտեգրման կարևորությունը։ Այսպիսով, սաղմի առաջացման յուրաքանչյուր փուլ ոչ միայն բջիջների քանակի ավելացում է, այլ ամբողջականության նոր վիճակ:
Ինտեգրումը բջիջների պոպուլյացիաների միավորումն է ավելի բարդ գործող համակարգերի` հյուսվածքների, օրգանների: Այն կարող է կոտրվել վիրուսների, բակտերիաների, ռենտգենյան ճառագայթների, հորմոնների և այլ գործոնների պատճառով: Այս դեպքերում կենսաբանական համակարգը դուրս է գալիս վերահսկողությունից, ինչը կարող է առաջացնել չարորակ ուռուցքների և այլ պաթոլոգիաների զարգացում։
Մորֆոֆունկցիոնալ և գենետիկական տարբերությունները, որոնք առաջացել են ֆիլոգենեզի գործընթացում, թույլ են տվել բջիջներին և ոչ բջջային կառուցվածքներին միավորվել այսպես կոչված հյուսվածաբանական հյուսվածքների մեջ։
Հյուսվածքը բջիջների և ոչ բջջային կառուցվածքների պատմականորեն հաստատված համակարգ է, որը բնութագրվում է ընդհանուր կառուցվածքով, գործառույթով և ծագմամբ:
Հյուսվածքների չորս հիմնական տեսակ կա՝ էպիթելային, շարակցական կամ օժանդակ-տրոֆիկ, մկանային և նյարդային։ Կան նաև այլ դասակարգումներ.
2. Էպիթելային հյուսվածքները մարմնին հաղորդակցում են արտաքին միջավայրի հետ։ Կատարում են ներքին և գեղձային (սեկրետորային) ֆունկցիաներ։ Էպիթելը գտնվում է մաշկի մեջ, գծում է բոլոր ներքին օրգանների լորձաթաղանթները; այն ունի կլանման, արտազատման գործառույթներ։ Մարմնի գեղձերի մեծ մասը կառուցված է էպիթելային հյուսվածքից։
Բոլոր սաղմնային շերտերը մասնակցում են էպիթելային հյուսվածքի զարգացմանը։
Բոլոր էպիթելիաները կառուցված են էպիթելային բջիջներից՝ էպիթելիոցիտներից: Դեսմոսոմների, փակող ժապավենների, սոսնձող ժապավենների օգնությամբ և միաձուլման միջոցով ամուր միանալով միմյանց՝ էպիթելիոցիտները կազմում են բջջային շերտ, որը գործում և վերականգնվում է: Որպես կանոն, շերտերը տեղակայված են նկուղային թաղանթի վրա, որն, իր հերթին, ընկած է չամրացված կապի հյուսվածքի վրա, որը սնուցում է էպիթելը (նկ. 7):
Էպիթելային հյուսվածքները բնութագրվում են բևեռային տարբերակմամբ, որը կրճատվում է էպիթելային շերտի այլ կառուցվածքի կամ շերտերի կամ էպիթելիոցիտների բևեռների: Օրինակ, գագաթային բևեռում պլազմոլեմման ձևավորում է ներծծող սահման կամ թարթիչավոր թարթիչներ, մինչդեռ միջուկը և օրգանելների մեծ մասը գտնվում են բազալ բևեռում:
Կախված գտնվելու վայրից և կատարվող ֆունկցիայից՝ առանձնանում են էպիթելի երկու տեսակ՝ ծածկված և գեղձային։
Ներքին էպիթելի ամենատարածված դասակարգումը հիմնված է բջիջների ձևի և էպիթելի շերտի շերտերի քանակի վրա, ուստի այն կոչվում է ձևաբանական:
3. Գաղտնիքներ արտադրող էպիթելը կոչվում է գեղձային, իսկ նրա բջիջները՝ սեկրետորային բջիջներ կամ սեկրետորային գեղձեր։ Գեղձերը կառուցված են արտազատող բջիջներից, որոնք կարող են նախագծվել որպես անկախ օրգան կամ միայն դրա մի մասն են։
Տարբերակել էնդոկրին և էկզոկրին գեղձերը: Մորֆոլոգիապես տարբերություն կա վերջինիս մոտ արտազատվող ծորանի առկայության հարցում։ Էկզոկրին գեղձերը կարող են լինել միաբջիջ կամ բազմաբջիջ: Օրինակ՝ գավաթային բջիջ պարզ սյունաձեւ եզրային էպիթելիում: Արտազատման ծորանի ճյուղավորման բնույթով առանձնանում են պարզ և բարդ։ Պարզ գեղձերը ունեն ոչ ճյուղավոր արտազատվող ծորան, իսկ բարդ գեղձերը՝ ճյուղավորվող։ Պարզ գեղձերի տերմինալային հատվածները ճյուղավորվում և չճյուղավորվում են, բարդ խցուկներում՝ ճյուղավորվում։
Ըստ տերմինալային հատվածների ձևի՝ էկզոկրին գեղձերը դասակարգվում են ալվեոլային, խողովակային և գլանային-ալվեոլային: Վերջնական հատվածի բջիջները կոչվում են գեղձի բջիջներ:
Ըստ սեկրեցիայի ձևավորման մեթոդի՝ գեղձերը բաժանվում են հոլոկրին, ապոկրին և մերոկրին։ Սրանք ստամոքսի համապատասխանաբար ճարպային, ապա քրտինքի և կաթնագեղձերն են։
Վերածնում. Integumentary էպիթելը զբաղեցնում է սահմանային դիրք: Նրանք հաճախ են վնասվում, հետևաբար դրանք բնութագրվում են վերականգնողական բարձր հզորությամբ։ Վերածնումն իրականացվում է հիմնականում միտոտիկ մեթոդով։ Էպիթելային շերտի բջիջները արագ մաշվում են, ծերանում և մահանում։ Դրանց վերականգնումը կոչվում է ֆիզիոլոգիական ռեգեներացիա։ Վնասվածքի պատճառով կորցրած էպիթելի բջիջների վերականգնումը կոչվում է ռեպարատիվ վերականգնում։
Միաշերտ էպիթելում բոլոր բջիջներն ունեն վերականգնողական հատկություն, բազմաշերտ էպիթելում՝ ցողունային բջիջներ։ Գեղձի էպիթելում, հոլոկրինային սեկրեցիայով, նկուղային թաղանթի վրա տեղակայված ցողունային բջիջներն ունեն այս հատկությունը։ Մերոկրին և ապոկրին գեղձերում էպիթելիոցիտների վերականգնումն ընթանում է հիմնականում ներբջջային ռեգեներացիայի միջոցով։

Բրինձ. 7. Էպիթելի տարբեր տեսակների սխեմա
A. Միաշերտ հարթ:
B. Մեկ շերտ խորանարդ.
B. Միաշերտ գլանաձեւ:
G. Բազմաշար գլանաձև թարթիչավոր:
D. Անցումային.
E. Բազմաշերտ հարթ չկերատինացված:
G. Բազմաշերտ հարթ կերատինացում:
Աջակցող-տրոֆիկ հյուսվածքներ. արյուն և ավիշ
1. Արյուն. Արյան բջիջներ.
3. Հեմոցիտոպոեզիա.
4. Սաղմնային հեմոցիտոպոեզիա:
Այս թեմայով մենք սկսում ենք մի խումբ հարակից հյուսվածքների ուսումնասիրությունը, որոնք կոչվում են կապող: Դրանք ներառում են՝ համապատասխան շարակցական հյուսվածք, արյան բջիջներ և արյունաստեղծ հյուսվածքներ, կմախքային հյուսվածքներ (աճառային և ոսկրային), հատուկ հատկություններով շարակցական հյուսվածքներ։
Հյուսվածքների վերոհիշյալ տեսակների միասնության դրսեւորումը նրանց ծագումն է ընդհանուր սաղմնային աղբյուրից՝ մեզենխիմից։
Մեզենխիմ - սաղմնային ցանցանման գործընթացային բջիջների մի շարք, որոնք լրացնում են բողբոջային շերտերի և օրգանների հիմքերի միջև եղած բացերը: Սաղմի մարմնում մեզենխիմը հիմնականում առաջանում է մեզոդերմի որոշակի հատվածների բջիջներից՝ դերմատոմներից, սկլերոտոմներից և սպլանխնոտոմներից։ Մեզենխիմային բջիջները արագորեն բաժանվում են միտոզով: Բազմաթիվ մեզենխիմալ ածանցյալներ առաջանում են նրա տարբեր մասերում՝ արյան կղզիներ իրենց էնդոթելիով և արյան բջիջներով, շարակցական հյուսվածքների և հարթ մկանային հյուսվածքի բջիջներով և այլն։
1. Ներանոթային արյուն՝ շարժական հյուսվածքային համակարգ՝ հեղուկ միջբջջային նյութով՝ պլազմա և ձևավորված տարրեր՝ էրիթրոցիտներ, լեյկոցիտներ և թրոմբոցիտներ։
Անընդհատ շրջանառվելով փակ շրջանառության համակարգում, արյունը միավորում է մարմնի բոլոր համակարգերի աշխատանքը և պահպանում է մարմնի ներքին միջավայրի բազմաթիվ ֆիզիոլոգիական ցուցանիշներ որոշակի մակարդակում, որն օպտիմալ է նյութափոխանակության գործընթացների համար: Արյունն օրգանիզմում կատարում է տարբեր կենսական գործառույթներ՝ շնչառական, տրոֆիկ, պաշտպանիչ, կարգավորող, արտազատող և այլն։
Չնայած արյան շարժունակությանը և փոփոխականությանը, նրա ցուցիչները ամեն պահի համապատասխանում են օրգանիզմի ֆունկցիոնալ վիճակին, ուստի արյան հետազոտությունը ախտորոշման կարևորագույն մեթոդներից է։
Պլազմա - արյան հեղուկ բաղադրիչ, պարունակում է 90-92% ջուր և 8-10% պինդ նյութեր, ներառյալ 9% օրգանական և 1% հանքային նյութեր: Արյան պլազմայի հիմնական օրգանական նյութերը սպիտակուցներն են (ալբումիններ, գլոբուլինների տարբեր ֆրակցիաներ և ֆիբրինոգեն)։ Իմունային սպիտակուցները (հակամարմինները), և դրանց մեծ մասը պարունակվում է գամմա գլոբուլինի ֆրակցիայում, կոչվում են իմունոգլոբուլիններ: Ալբոմիններն ապահովում են տարբեր նյութերի փոխանցում՝ ազատ ճարպաթթուներ, բիլիռուբին և այլն։ Ֆիբրինոգենը մասնակցում է արյան մակարդման գործընթացներին։
Էրիտրոցիտները արյան բջիջների հիմնական տեսակն են, քանի որ դրանք 500-1000 անգամ ավելի շատ են, քան լեյկոցիտները։ Խոշոր եղջերավոր անասունների արյան 1 մմ 3-ը պարունակում է 5,0-7,5 մլն, ձիերինը՝ 6-9 մլն, ոչխարներինը՝ 7-12 մլն, այծերինը՝ 12-18 մլն, խոզերինը՝ 6-7,5 մլն, հավերինը՝ 3-4 մլն կարմիր արյուն: բջիջները.
Զարգացման ընթացքում կորցնելով միջուկը՝ կաթնասունների հասուն էրիթրոցիտները միջուկից զուրկ բջիջներ են և ունեն երկգոգավոր սկավառակի ձև՝ 5-7 մկմ միջին շրջանագծի տրամագծով։ Ուղտի և լամայի արյան էրիթրոցիտները օվալ են։ Սկավառակի ձևը մեծացնում է էրիթրոցիտների ընդհանուր մակերեսը 1,64 անգամ։
Արյան կարմիր բջիջների քանակի և դրանց չափի միջև կա հակադարձ կապ:
Էրիտրոցիտները ծածկված են թաղանթով՝ պլազմոլեմմայով (6 նմ հաստությամբ), որը պարունակում է 44% լիպիդներ, 47% սպիտակուցներ և 7% ածխաջրեր։ Էրիտրոցիտների թաղանթը հեշտությամբ թափանցելի է գազերի, անիոնների և Na իոնների համար։
Էրիտրոցիտների ներքին կոլոիդային պարունակությունը 34%-ով բաղկացած է հեմոգլոբինից՝ եզակի բարդ գունավոր միացությունից՝ քրոմպրոտեինից, որի ոչ սպիտակուցային մասում (հեմում) կա գունավոր երկաթ, որն ունակ է թթվածնի մոլեկուլի հետ հատուկ փխրուն կապեր ստեղծել։ Հեմոգլոբինի շնորհիվ է իրականացվում էրիթրոցիտների շնչառական ֆունկցիան։ Օքսիհեմոգլոբին \u003d հեմոգլոբին + O2:
Էրիտրոցիտներում հեմոգլոբինի առկայությունը առաջացնում է նրանց ընդգծված օքսիֆիլիա՝ ըստ Ռոմանովսկի-Գիմսայի (էոզին + լազուր II) արյան քսուք ներկելիս: Էրիտրոցիտները կարմիր ներկված են էոզինով։ Անեմիայի որոշ ձևերի դեպքում մեծանում է էրիթրոցիտների կենտրոնական գունատ գույնի մասը՝ հիպոքրոմային էրիթրոցիտները։ Սուպրավիտալ արյան ներկման դեպքում փայլուն կրեսիլ կապույտով կարելի է հայտնաբերել էրիթրոցիտների երիտասարդ ձևեր, որոնք պարունակում են հատիկավոր ցանցային կառուցվածքներ: Նման բջիջները կոչվում են ռետիկուլոցիտներ, դրանք հասուն էրիթրոցիտների անմիջական պրեկուրսորներն են։ Ռետիկուլոցիտների հաշվարկը օգտագործվում է կարմիր արյան բջիջների ձևավորման արագության մասին տեղեկատվություն ստանալու համար:
Էրիտրոցիտների կյանքի տևողությունը 100-130 օր է (ճագարների մոտ՝ 45-60 օր): Էրիտրոցիտները կարող են դիմակայել տարբեր կործանարար ազդեցություններին՝ օսմոտիկ, մեխանիկական և այլն: Շրջակա միջավայրում աղերի կոնցենտրացիայի փոփոխության դեպքում էրիթրոցիտների թաղանթը դադարում է պահպանել հեմոգլոբինը, և այն մտնում է շրջակա հեղուկ՝ հեմոլիզի ֆենոմենը: Հեմոգլոբինի արտազատումը օրգանիզմում կարող է առաջանալ օձի թույնի, տոքսինների ազդեցության տակ։ Հեմոլիզը զարգանում է նաև անհամատեղելի արյան խմբերի փոխներարկման ժամանակ։ Կենդանիների արյան մեջ հեղուկներ ներմուծելիս գործնականում կարևոր է վերահսկել, որ ներարկվող լուծույթը իզոտոնիկ է:
Էիթրոցիտները համեմատաբար բարձր խտություն ունեն՝ համեմատած պլազմայի և լեյկոցիտների հետ։ Եթե արյունը մշակվում է հակակոագուլանտներով և տեղադրվում է անոթի մեջ, ապա նշվում է էրիթրոցիտների նստվածք։ Տարբեր տարիքի, սեռի և տեսակների կենդանիների մոտ էրիթրոցիտների նստվածքի արագությունը (ESR) նույնը չէ: Ձիերի մեջ բարձր ESR և, ընդհակառակը, անասունների ցածր մակարդակ: ESR-ն ունի ախտորոշիչ և պրոգնոստիկ արժեք:
Լեյկոցիտները անոթային արյան բջիջներ են, որոնք բազմազան են մորֆոլոգիական առանձնահատկություններով և գործառույթներով: Կենդանիների մարմնում նրանք կատարում են մի շարք գործառույթներ, որոնք հիմնականում ուղղված են պաշտպանելու մարմինը օտար ազդեցություններից ֆագոցիտային գործունեության, հումորալ և բջջային անձեռնմխելիության ձևավորմանը մասնակցելու, ինչպես նաև հյուսվածքների վնասման վերականգնման գործընթացներին: Անասունների 1 մմ3 արյան մեջ կա 4,5-12 հազար, ձիերինը՝ 7-12 հազար, ոչխարներինը՝ 6-14 հազար, խոզերինը՝ 8-16 հազար, հավերինը՝ 20-40 հազար:Աճ է լեյկոցիտների քանակում՝ լեյկոցիտոզը, բնորոշ հատկանիշ է բազմաթիվ պաթոլոգիական պրոցեսների համար։
Ձևավորվելով արյունաստեղծ օրգաններում և մտնելով արյուն՝ լեյկոցիտները միայն կարճ ժամանակ են մնում անոթային անկողնում, այնուհետև գաղթում են շրջակա անոթային շարակցական հյուսվածք և օրգաններ, որտեղ կատարում են իրենց հիմնական գործառույթը։
Լեյկոցիտների առանձնահատկությունն այն է, որ նրանք ունեն շարժունակություն՝ առաջացած պսեւդոպոդիայի շնորհիվ։ Լեյկոցիտներում առանձնանում են միջուկը և ցիտոպլազմը, որոնք պարունակում են տարբեր օրգանելներ և ներդիրներ։ Լեյկոցիտների դասակարգումը հիմնված է ներկերով ներկելու ունակության և հատիկավորության վրա:
Հատիկավոր լեյկոցիտներ (գրանուլոցիտներ)՝ նեյտրոֆիլներ (25-70%), էոզինոֆիլներ (2-12%), բազոֆիլներ (0,5-2%)։
Ոչ հատիկավոր լեյկոցիտներ (ագրանուլոցիտներ)՝ լիմֆոցիտներ (40-65) և մոնոցիտներ (1-8%)։
Լեյկոցիտների առանձին տեսակների միջև որոշակի տոկոսային հարաբերակցությունը կոչվում է լեյկոցիտային բանաձև՝ լեյկոգրամ:
Թարախային-բորբոքային պրոցեսներին բնորոշ է լեյկոգրամում նեյտրոֆիլների տոկոսի ավելացումը։ Հասուն նեյտրոֆիլներում միջուկը բաղկացած է մի քանի հատվածներից, որոնք միացված են բարակ կամուրջներով։
Բազոֆիլների մակերեսին կան հատուկ ընկալիչներ, որոնք կապում են E իմունոգոլոբուլինները: Նրանք մասնակցում են ալերգիկ տիպի իմունոլոգիական ռեակցիաներին:
Արյան մեջ շրջանառվող մոնոցիտները հյուսվածքների և օրգանների մակրոֆագների պրեկուրսորներ են: Անոթային արյան մեջ մնալուց հետո (12-36 ժամ) մոնոցիտները մազանոթների և վենուլների էնդոթելիով գաղթում են հյուսվածքներ և վերածվում շարժական մակրոֆագների։
Լիմֆոցիտները ամենակարևոր բջիջներն են, որոնք ներգրավված են մարմնի տարբեր իմունոլոգիական ռեակցիաներում: Լիմֆայում հայտնաբերվում են մեծ քանակությամբ լիմֆոցիտներ:
Լիմֆոցիտների երկու հիմնական դաս կա՝ T- և B-լիմֆոցիտներ: Առաջինը զարգանում է ոսկրածուծի բջիջներից՝ տիմուսի լոբուլների կեղևային մասում: Պլազմալեմայում ունեն հակագենային մարկերներ և բազմաթիվ ընկալիչներ, որոնց օգնությամբ ճանաչվում են օտար անտիգենները և իմունային համալիրները։
B-լիմֆոցիտները ձևավորվում են ցողունային պրեկուրսորներից Fabricius-ի (Բուրսա) բուրսայում: Նրանց զարգացման վայրը համարվում է ոսկրածուծի միելոիդ հյուսվածք:
T-լիմֆոցիտային համակարգում գոյություն ունեն էֆեկտորային բջիջների երեք հիմնական ենթապոպուլյացիաներ՝ T-մարդասպաններ (ցիտոտոքսիկ լիմֆոցիտներ), T-helpers (օգնականներ) և T-suppressors (դեպրեսորներ): B-լիմֆոցիտների էֆեկտոր բջիջները պլազմաբլաստներն են և հասուն պլազմային բջիջները, որոնք ունակ են ավելացել է գումարըարտադրում է իմունոգոլոբուլիններ.
Թրոմբոցիտները կաթնասունների անոթային արյան ոչ միջուկային տարրեր են։ Սրանք կարմիր ոսկրածուծի մեգակարիոցիտների ցիտոպլազմիկ բեկորներ են: Նրանց արյան 1 մմ3-ում կա 250-350 հազար թրոմբոցիտ։ Թռչունների մոտ բջիջները, որոնք գործում են նույն կերպ, կոչվում են թրոմբոցիտներ:
Արյան թրոմբոցիտները ամենակարևոր գիտելիքներն ունեն արյունահոսությունը դադարեցնելու հիմնական փուլերի՝ հեմոստազի ապահովման հարցում։
2. Լիմֆ - գրեթե թափանցիկ դեղնավուն հեղուկ, որը գտնվում է ավշային մազանոթների և անոթների խոռոչում: Դրա ձևավորումը պայմանավորված է անցումով բաղկացուցիչ մասերարյան պլազմա արյան մազանոթներից մինչև հյուսվածքային հեղուկ: Լիմֆի ձևավորման ժամանակ էական նշանակություն ունեն արյան և հյուսվածքային հեղուկի հիդրոստատիկ և օսմոտիկ ճնշման հարաբերությունները, արյան մազանոթների պատերի թափանցելիությունը և այլն։
Լիմֆը բաղկացած է հեղուկ մասից՝ լիմֆոպլազմից և ձևավորված տարրերից։ Լիմֆոպլազման արյան պլազմայից տարբերվում է սպիտակուցների ավելի ցածր պարունակությամբ։ Լիմֆը պարունակում է ֆիբրինոգեն, ուստի այն նաև ունակ է մակարդվել։ Լիմֆի հիմնական ձևավորված տարրերը լիմֆոցիտներն են: Լիմֆի բաղադրությունը ավշային համակարգի տարբեր անոթներում նույնը չէ։ Տարբերում են ծայրամասային (ավշահանգույցներից առաջ), միջանկյալ (ավշահանգույցներից հետո) և կենտրոնական (կրծքավանդակի և աջ ավշային խողովակների լիմֆա), որոնք ամենահարուստն են բջջային տարրերով։
3. Արյունաստեղծումը (հեմոցիտոպոեզը) բջջային հաջորդական փոխակերպումների բազմաստիճան գործընթաց է, որը հանգեցնում է հասուն ծայրամասային անոթային արյան բջիջների ձևավորմանը:
Կենդանիների հետսեմբրիոնային շրջանում արյան բջիջների զարգացումն իրականացվում է երկու մասնագիտացված ինտենսիվ վերականգնվող հյուսվածքներում՝ միելոիդ և լիմֆոիդ:
Ներկայումս ամենաճանաչվածը արյունաստեղծման սխեման է, որն առաջարկել է Ի.Լ. Չերտկովը և Ա.Ի. Վորոբյովը (1981 թ.), ըստ որի ամբողջ հեմոցիտոպոեզը բաժանվում է 6 փուլերի (նկ. 8):
Արյան բոլոր բջիջների նախահայրը (ըստ Ա.Ա. Մաքսիմովի) պլյուրիպոտենտ ցողունային բջիջ է (գաղութ ձևավորող միավոր փայծաղում և CFU-ներում): Մեծահասակների մարմնում ցողունային բջիջների ամենամեծ քանակությունը գտնվում է կարմիր ոսկրածուծում (մոտ 50 ցողունային բջիջ 100000 ոսկրածուծի բջիջից), որտեղից նրանք գաղթում են դեպի տիմուս՝ փայծաղ։
Կարմիր ոսկրածուծում էրիթրոցիտների զարգացումը (էրիթրոցիտոպոեզը) ընթանում է ըստ սխեմայի՝ ցողունային բջիջ (SC) - կիսատև ցողունային բջիջներ (CFU - GEMM, CFU - GE, CFU - MGCE) - էրիթրոպոեզի միատև պրեկուրսորներ (PFU - E): , CFU - E) - erythroblast - pronormocyte - basophilic normocyte - polychromatophilic normocyte - oxyphilic normocyte - reticulocyte - erythrocyte.
Գրանուլոցիտների զարգացում. կարմիր ոսկրածուծի ցողունային բջիջ, կիսա-ցողուն (CFU - GEMM, CFU - GM, CFU - GE), միաուժ պրեկուրսորներ (CFU - B, CFU - Eo, CFU - Gn), որոնք անցնում են փուլերով. Բջջային ճանաչելի ձևերը վերածվում են հասուն հատվածավորված միջուկային գրանուլոցիտների երեք սորտերի՝ նեյտրոֆիլների, էոզինոֆիլների և բազոֆիլների:
Լիմֆոցիտների զարգացումը արյունաստեղծ ցողունային բջիջների տարբերակման ամենաբարդ գործընթացներից մեկն է:
Տարբեր օրգանների մասնակցությամբ փուլ առ փուլ իրականացվում է երկու սերտ կապված բջջային գծերի՝ T- և B-լիմֆոցիտների ձևավորումը։
Թրոմբոցիտների զարգացումը տեղի է ունենում կարմիր ոսկրածուծում և կապված է դրանում հատուկ հսկա բջիջների՝ մեգակարիոցիտների զարգացման հետ։ Մեգակարիոցիտոպոեզը բաղկացած է հետևյալ փուլերից՝ SC - կիսագնդային բջիջներ (CFU - GEMM և CFU - MGCE) - unipotent պրեկուրսորներ, (CFU - MHC) - megakaryoblast - promegakaryocyte - megakaryocyte:
4. Օնտոգենեզի ամենավաղ փուլերում արյան բջիջները ձևավորվում են սաղմից դուրս՝ դեղնուցի պարկի մեզենխիմում, որտեղ ձևավորվում են կլաստերներ՝ արյան կղզիներ։ Կղզիների կենտրոնական բջիջները կլորացվում են և վերածվում արյունաստեղծ ցողունային բջիջների։ Կղզիների ծայրամասային բջիջները ձգվում են շերտերով, փոխկապակցված բջիջներով և կազմում են առաջնային արյան անոթների էնդոթելի լորձաթաղանթը (դեղնապարկի անոթը)։ Ցողունային բջիջների մի մասը վերածվում է խոշոր բազոֆիլային բլաստ բջիջների՝ առաջնային արյան բջիջների։ Այս բջիջների մեծ մասը, ինտենսիվ բազմանալով, ավելի ու ավելի են ներկվում թթվային ներկերով։ Դա տեղի է ունենում ցիտոպլազմայում հեմոգլոբինի սինթեզի և կուտակման, իսկ միջուկում խտացված քրոմատինի հետ կապված: Այս բջիջները կոչվում են առաջնային էրիթրոբլաստներ: Որոշ առաջնային էրիթրոբլաստներում միջուկը քայքայվում և անհետանում է: Միջուկային և ոչ միջուկային առաջնային էրիթրոցիտների առաջացող սերունդը չափսերով բազմազան է, սակայն ամենատարածվածը խոշոր բջիջներն են՝ մեգալոբլաստները և մեգալոցիտները։ Սաղմնային շրջանին բնորոշ է արյունաստեղծության մեգալոբլաստիկ տեսակը։
Արյան առաջնային բջիջների մի մասը վերածվում է երկրորդային էրիթրոցիտների պոպուլյացիայի, իսկ անոթներից դուրս զարգանում են փոքր քանակությամբ գրանուլոցիտներ՝ նեյտրոֆիլներ և էոզինոֆիլներ, այսինքն՝ առաջանում է միելոպոեզ։
Ցողունային բջիջները, որոնք առաջանում են դեղնուցի պարկի մեջ, արյունով տեղափոխվում են մարմնի օրգաններ։ Լյարդը դնելուց հետո այն դառնում է արյունաստեղծման ունիվերսալ օրգան (զարգանում են երկրորդային էրիթրոցիտներ, հատիկավոր լեյկոցիտներ և մեգակարիոցիտներ)։ Ներարգանդային շրջանի վերջում լյարդում արյունաստեղծումը դադարում է։
Սաղմնային զարգացման 7-8 շաբաթվա ընթացքում (խոշոր եղջերավոր անասունների մոտ) թիմուսի լիմֆոցիտները և դրանից գաղթող T-լիմֆոցիտները տարբերվում են զարգացող թիմուսի ցողունային բջիջներից։ Վերջիններս բնակվում են փայծաղի և ավշային հանգույցների T գոտիներում։ Իր զարգացման սկզբում փայծաղը նաև օրգան է, որտեղ ձևավորվում են արյան բոլոր տեսակի բջիջները։
Կենդանիների սաղմնային զարգացման վերջին փուլերում հիմնական արյունաստեղծ գործառույթները սկսում են իրականացնել կարմիր ոսկրածուծը. արտադրում է էրիթրոցիտներ, գրանուլոցիտներ, թրոմբոցիտներ, լիմֆոցիտների մի մասը (V-l): Հետսամբրիոնային շրջանում կարմիր ոսկրածուծը դառնում է համընդհանուր արյունաստեղծման օրգան։
Սաղմնային էրիթրոցիտոպոեզի ժամանակ տեղի է ունենում էրիթրոցիտների սերունդների փոփոխման բնորոշ գործընթաց, որոնք տարբերվում են ձևաբանությամբ և ձևավորված հեմոգլոբինի տեսակով։ Առաջնային էրիթրոցիտների պոպուլյացիան կազմում է հեմոգլոբինի սաղմնային տեսակը (Hb - F): Հետագա փուլերում լյարդի և փայծաղի էրիթրոցիտները պարունակում են պտղի (պտղի) տեսակի հեմոգլոբին (Hb-G): Կարմիր ոսկրածուծում ձևավորվում են էրիթրոցիտների վերջնական տեսակը երրորդ տեսակի հեմոգլոբինով (Hb-A և Hb-A 2): Հեմոգլոբինների տարբեր տեսակներ տարբերվում են սպիտակուցային մասի ամինաթթուների կազմով։
բջիջների սաղմի գենեզի հյուսվածքների հիստոլոգիա բջջաբանություն
Միակցիչ հյուսվածքը ճիշտ է
1. Չամրացված և խիտ շարակցական հյուսվածք:
2. Հատուկ հատկություններով շարակցական հյուսվածք՝ ցանցանման, ճարպային, պիգմենտային։
1. Կենդանիների օրգանիզմում տարածված հյուսվածքներ միջբջջային նյութի մանրաթելերի բարձր զարգացած համակարգով, որի շնորհիվ այդ հյուսվածքները կատարում են բազմակողմանի մեխանիկական և ձևավորող գործառույթներ. , ձևավորել պարկուճներ, կապաններ, ֆասիա, ջլեր:
Կախված միջբջջային նյութի բաղադրիչների՝ մանրաթելերի և հիմնական նյութի քանակական հարաբերակցությունից և ըստ մանրաթելերի տեսակի՝ առանձնանում են երեք տեսակի շարակցական հյուսվածք՝ չամրացված շարակցական հյուսվածք, խիտ շարակցական հյուսվածք և ցանցանման հյուսվածք։
Հիմնական բջիջները, որոնք ստեղծում են նյութեր, որոնք անհրաժեշտ են չամրացված և խիտ շարակցական հյուսվածքում մանրաթելեր կառուցելու համար, ֆիբրոբլաստներն են, ցանցային հյուսվածքում՝ ցանցանման բջիջները։ Չամրացված շարակցական հյուսվածքը բնութագրվում է բջջային կազմի առանձնապես մեծ բազմազանությամբ:
Չամրացված շարակցական հյուսվածքը ամենատարածվածն է: Այն ուղեկցում է բոլոր արյան և ավշային անոթներին, օրգանների ներսում ձևավորում է բազմաթիվ շերտեր և այլն: Այն բաղկացած է մի շարք բջիջներից, հիմնական նյութից և կոլագենի և առաձգական մանրաթելերի համակարգից: Այս հյուսվածքի բաղադրության մեջ առանձնանում են ավելի շատ նստակյաց բջիջներ (ֆիբրոբլաստներ՝ ֆիբրոցիտներ, լիպոցիտներ), շարժական (հիստիոցիտներ՝ մակրոֆագներ, հյուսվածքային բազոֆիլներ, պլազմոցիտներ) - Նկ. 9։
Այս կապ հյուսվածքի հիմնական գործառույթներն են տրոֆիկ, պաշտպանիչ և պլաստիկ:
Բջիջների տեսակները. Ադվենցիալ բջիջները վատ տարբերակված են, ունակ են միտոտիկ բաժանման և վերածվելու ֆիբրոբլաստների, միոֆիբրոբլաստների և լիպոցիտների: Ֆիբրոբլաստները հիմնական բջիջներն են, որոնք անմիջականորեն մասնակցում են միջբջջային կառուցվածքների ձևավորմանը: Սաղմի զարգացման ընթացքում ֆիբրոբլաստները առաջանում են անմիջապես մեզենխիմային բջիջներից: Կան երեք տեսակի ֆիբրոբլաստներ՝ վատ տարբերակված (գործառույթը՝ գլիկոզամինոգլիկանների սինթեզ և սեկրեցիա); հասուն (գործառույթը՝ պրոկոլագենի, պրոելաստինի, ֆերմենտային սպիտակուցների և գլիկոզամինոգլիկանների սինթեզ, հատկապես՝ կոլագենի մանրաթելերի սպիտակուցային սինթեզ); միոֆիբրոբլաստներ, որոնք նպաստում են վերքերի փակմանը: Ֆիբրոցիտները կորցնում են բաժանվելու ունակությունը, նվազեցնում սինթետիկ ակտիվությունը։ Հիստիոցիտները (մակրոֆագները) պատկանում են միամիջուկային ֆագոցիտների համակարգին (MPS): Այս համակարգը կքննարկվի հաջորդ դասախոսության ժամանակ: Հյուսվածքային բազոֆիլները (լաբրոցիտներ, մաստ բջիջներ), որոնք տեղակայված են փոքր արյան անոթների մոտ, դրանք առաջին բջիջներից են, որոնք արձագանքում են արյունից անտիգենների ներթափանցմանը:
Պլազմոցիդներ - ֆունկցիոնալ - հումորալ տիպի իմունոլոգիական ռեակցիաների էֆեկտորային բջիջներ: Սրանք մարմնի բարձր մասնագիտացված բջիջներ են, որոնք սինթեզում և արտազատում են տարբեր հակամարմինների (իմունոգլոբուլինների) հիմնական մասը:
Դրա զգալի մասն է կազմում չամրացված շարակցական հյուսվածքի միջբջջային նյութը։ Այն ներկայացված է կոլագենի և առաձգական մանրաթելերով և հիմնական (ամորֆ) նյութով։
Ամորֆ նյութ - շարակցական հյուսվածքի բջիջների (հիմնականում ֆիբրոբլաստների) սինթեզի և արյունից նյութերի ընդունման արդյունք, թափանցիկ, թեթևակի դեղնավուն, որը կարող է փոխել իր հետևողականությունը, ինչը զգալիորեն ազդում է դրա հատկությունների վրա:
Այն բաղկացած է գլիկոզամինոգլիկաններից (պոլիսաքարիդներից), պրոտեոգլիկաններից, գլիկոպրոտեիններից, ջրից և անօրգանական աղերից։ Այս համալիրի ամենակարևոր քիմիական բարձր պոլիմերային նյութը գլիկոզամինոգլիկանների չսուլֆատացված բազմազանությունն է՝ հիալուրոնաթթուն:
Կոլագենային մանրաթելերը կազմված են տրոպոկոլագենի սպիտակուցի մոլեկուլներից ձևավորված մանրաթելերից։ Վերջիններս առանձնահատուկ մոնոմերներ են։ Ֆիբրիլների առաջացումը երկայնական և լայնակի ուղղություններով մոնոմերների բնորոշ խմբավորման արդյունք է։
Կախված ամինաթթուների կազմից և շղթաների ձևից, որոնք միավորվում են եռակի պարույրի մեջ, կան կոլագենի չորս հիմնական տեսակներ, որոնք ունեն տարբեր տեղայնացում մարմնում: I տիպի կոլագենը հայտնաբերված է մաշկի, ջլերի և ոսկորների միացնող հյուսվածքում: Կոլագեն II տիպ - հիալինային և մանրաթելային աճառներում: Կոլագեն II? տեսակ - սաղմերի մաշկի, արյան անոթների պատի, կապանների մեջ: Կոլագեն տիպի IV - նկուղային թաղանթներում:
Կոլագենի մանրաթելերի ձևավորման երկու եղանակ կա՝ ներբջջային և արտաբջջային սինթեզ։
Էլաստիկ մանրաթելերը միատարր թելեր են, որոնք ցանց են կազմում։ Մի միավորեք փաթեթների մեջ, ունենան ցածր ուժ: Կա ավելի թափանցիկ ամորֆ կենտրոնական մաս՝ կազմված էլաստին սպիտակուցից, և ծայրամասային մասը՝ գլիկոպրոտեինային բնույթի միկրոֆիբրիլներից՝ խողովակների ձևով։ Էլաստիկ մանրաթելերը ձևավորվում են ֆիբրոբլաստների սինթետիկ և սեկրետորային ֆունկցիայի շնորհիվ։ Ենթադրվում է, որ սկզբում, ֆիբրոբլաստների անմիջական հարևանությամբ, ձևավորվում է միկրոֆիբրիլների շրջանակ, այնուհետև ուժեղանում է էլաստինի պրեկուրսորից՝ պրոելաստինից, ամորֆ մասի ձևավորումը։ Պրոելաստինի մոլեկուլները ֆերմենտների ազդեցությամբ կրճատվում են և վերածվում տրոպոելաստինի մոլեկուլների։ Վերջիններս էլաստինի առաջացման ժամանակ փոխկապակցված են դեզմոզինի օգնությամբ, որը բացակայում է այլ սպիտակուցներում։ Էլաստիկ մանրաթելերը գերակշռում են օքսիպիտալ-արգանդի վզիկի կապում, որովայնի դեղին ֆասիայում։
Խիտ կապ հյուսվածք: Այս հյուսվածքը բնութագրվում է հիմնական նյութի և բջիջների նկատմամբ մանրաթելերի քանակական գերակշռությամբ: Կախված մանրաթելերի և ներքևից ձևավորված ցանցերի հարաբերական դիրքից՝ առանձնանում են խիտ շարակցական հյուսվածքի երկու հիմնական տեսակ՝ չձևավորված (դերմիս) և ձևավորված (ջլեր, ջլեր)։
2. Ցանցային հյուսվածքը բաղկացած է պրոցեսային ցանցային բջիջներից և ցանցաթելերից (նկ. 10): Ցանցային հյուսվածքը ձևավորում է արյունաստեղծ օրգանների ստրոման, որտեղ մակրոֆագերի հետ համատեղ ստեղծում է միկրոմիջավայր, որն ապահովում է արյան տարբեր բջիջների վերարտադրությունը, տարբերակումը և միգրացիան:
Ցանցային բջիջները զարգանում են մեզենխիմոցիտներից և նման են ֆիբրոբլաստներին, խոնդրոբլաստներին և այլն։ Դրանք պարունակում են տարբեր տրամագծերի մանրաթելեր՝ պարփակված միջֆիբրիլային նյութի մեջ։ Ֆիբրիլները կազմված են III տիպի կոլագենից։
Ճարպային հյուսվածքը կազմված է ճարպային բջիջներից (լիպոցիտներ): Վերջիններս մասնագիտացված են ցիտոպլազմայում պահեստային լիպիդների, հիմնականում՝ տրիգլիցերիդների սինթեզման և կուտակման գործում։ Լիպոցիտները լայնորեն տարածված են չամրացված շարակցական հյուսվածքում: Էմբրիոգենեզում ճարպային բջիջները առաջանում են մեզենխիմալ բջիջներից։
Հետսեմբրիոնային շրջանում նոր ճարպային բջիջների ձևավորման պրեկուրսորները արյան մազանոթներին ուղեկցող ադվենցիալ բջիջներն են:
Կան երկու տեսակի լիպոցիտներ և իրականում երկու տեսակի ճարպային հյուսվածք՝ սպիտակ և շագանակագույն: Սպիտակ ճարպային հյուսվածքը կենդանիների մարմնում պարունակվում է տարբեր կերպ՝ կախված տեսակից և ցեղից: Այն շատ է ճարպային պահեստներում: Տարբեր տեսակների, ցեղատեսակների, սեռի, տարիքի, գիրության կենդանիների օրգանիզմում դրա ընդհանուր քանակը տատանվում է ճարպային զանգվածի 1-30%-ի սահմաններում։ Ճարպը՝ որպես էներգիայի աղբյուր (1 գ ճարպ = 39 կՋ), ջրի պահեստ, շոկի կլանիչ։

Բրինձ. 11. Սպիտակ ճարպային հյուսվածքի կառուցվածքը (սխեմա ըստ Յու.Ի. Աֆանասիևի)
Ա - ճարպային բջիջներ հեռացված ճարպով թեթև օպտիկական մանրադիտակով; B - ճարպային բջիջների ուլտրամիկրոսկոպիկ կառուցվածքը: 1 - ճարպային բջիջի միջուկը; 2 - լիպիդների մեծ կաթիլներ; 3 - նյարդային մանրաթելեր; 4 - hemocapillaries; 5 - mitochondria.

Բրինձ. 12. Շագանակագույն ճարպային հյուսվածքի կառուցվածքը (սխեման ըստ Յու.Ի. Աֆանասիևի)
Ա - ճարպային բջիջներ հեռացված ճարպով թեթև օպտիկական մանրադիտակով; B - ճարպային բջիջների ուլտրամիկրոսկոպիկ կառուցվածքը: 1 - adipocyte կորիզ; 2 - նուրբ բաժանված լիպիդներ; 3 - բազմաթիվ միտոքոնդրիաներ; 4 - hemocapillaries; 5 - նյարդային մանրաթել.
Դարչնագույն ճարպային հյուսվածքը զգալի քանակությամբ հանդիպում է կրծողների և ձմեռային կենդանիների մոտ. ինչպես նաև այլ տեսակների նորածինների մոտ։ Բջիջները, օքսիդացած, ձևավորում են ջերմություն, որն անցնում է ջերմակարգավորման։
Պիգմենտային բջիջները (պիգմենտոցիտները) ունեն բազմաթիվ մուգ շագանակագույն կամ սև պիգմենտային հատիկներ ցիտոպլազմայի մելանինի խմբից:
Իմունային համակարգը և բջջային փոխազդեցությունները իմունային պատասխաններում
1. Հակագենների և հակամարմինների հայեցակարգը, դրանց տեսակները:
2 Բջջային և հումորալ իմունիտետի հայեցակարգը:
3 T- և B-լիմֆոցիտների ծագումը և փոխազդեցությունը:
4 Մակրոֆագների մոնոմիջուկային համակարգ.
1. Արդյունաբերական անասնաբուծության մեջ անասունների համակենտրոնացման և ինտենսիվ շահագործման, տեխնածին և այլ բնապահպանական գործոնների սթրեսային ազդեցության պայմաններում կենդանիների, հատկապես երիտասարդ կենդանիների հիվանդությունների կանխարգելման դերը, որոնք առաջանում են վարակիչ և ոչ վարակիչ բնույթի տարբեր գործակալների ազդեցության հետևանքով. , օրգանիզմի բնական պաշտպանիչ կարողությունների նվազման ֆոնին զգալիորեն ավելանում է։
Սրա շնորհիվ մեծ նշանակությունձեռք է բերում կենդանիների ֆիզիոլոգիական և իմունոլոգիական վիճակի վերահսկման խնդիր՝ նրանց ընդհանուր և հատուկ դիմադրությունը ժամանակին բարձրացնելու նպատակով (Tsymbal A.M., Konarzhevsky K.E. et al., 1984):
Իմունիտետը (իմունիտատիս՝ ինչ-որ բանից ազատում) մարմնի պաշտպանությունն է գենետիկորեն օտար ամեն ինչից՝ մանրէներից, վիրուսներից, այլմոլորակային բջիջներից։ կամ գենետիկորեն ձևափոխված սեփական բջիջները:
Իմունային համակարգը միավորում է օրգաններն ու հյուսվածքները, որոնցում ձևավորվում և փոխազդում են բջիջները՝ իմունոցիտները, որոնք կատարում են գենետիկորեն օտար նյութերի (անտիգենների) ճանաչման և հատուկ ռեակցիա իրականացնելու գործառույթը:
Հակամարմինները բարդ սպիտակուցներ են, որոնք հայտնաբերված են կենդանիների արյան պլազմայի իմունոգոլոբուլինային ֆրակցիայում, որոնք սինթեզվում են պլազմային բջիջների կողմից տարբեր անտիգենների ազդեցության տակ: Ուսումնասիրվել են իմունոգոլոբուլինների մի քանի դասեր (Y, M, A, E, D):
Հակագենի հետ առաջին հանդիպումից հետո (առաջնային պատասխան) լիմֆոցիտները գրգռվում են և փոխակերպվում են պայթյունի ձևերի, որոնք ունակ են բազմանալ և տարբերվել իմունոցիտների: Տարբերակումը հանգեցնում է երկու տեսակի բջիջների՝ էֆեկտորային և հիշողության բջիջների առաջացմանը: Առաջիններն ուղղակիորեն մասնակցում են օտար նյութերի վերացմանը: Էֆեկտոր բջիջները ներառում են ակտիվացված լիմֆոցիտներ և պլազմային բջիջներ: Հիշողության բջիջները լիմֆոցիտներ են, որոնք վերադառնում են ոչ ակտիվ վիճակի, բայց կրում են տեղեկատվություն (հիշողություն) կոնկրետ անտիգենի հետ հանդիպման մասին։ Այս անտիգենի կրկնակի ներդրմամբ նրանք կարողանում են արագ իմունային պատասխան ապահովել (երկրորդային պատասխան) լիմֆոցիտների ավելացման և իմունոցիտների ձևավորման շնորհիվ:
2. Կախված անտիգենների ոչնչացման մեխանիզմից՝ առանձնանում են բջջային իմունիտետը և հումորալ իմունիտետը։
Բջջային իմունիտետում էֆեկտոր (շարժիչ) բջիջները ցիտոտոքսիկ T-լիմֆոցիտներ են կամ սպանող լիմֆոցիտներ (մարդասպաններ), որոնք ուղղակիորեն մասնակցում են այլ օրգանների կամ սեփական պաթոլոգիական բջիջների օտար բջիջների ոչնչացմանը և արտազատում լիտիկ նյութեր: .
Հումորալ իմունիտետում էֆեկտոր բջիջները պլազմային բջիջներ են, որոնք սինթեզում և արյան մեջ հակամարմիններ են արտազատում:
Մարդկանց և կենդանիների բջջային և հումորալ իմունիտետի ձևավորման գործում կարևոր դեր են խաղում լիմֆոիդ հյուսվածքի բջջային տարրերը, մասնավորապես T- և B-լիմֆոցիտները: Անասունների արյան մեջ այդ բջիջների պոպուլյացիայի մասին տեղեկությունները սակավ են: Ըստ Կորչանի Ն.Ի. (1984), հորթերը ծնվում են համեմատաբար հասուն B-լիմֆոցիտային համակարգով և թերզարգացած B-լիմֆոցիտային համակարգով և այս բջիջների միջև կարգավորիչ հարաբերություններով: Միայն կյանքի 10-15-րդ օրը այդ բջջային համակարգերի ցուցանիշները մոտենում են չափահաս կենդանիների ցուցանիշներին:
Հասուն կենդանու օրգանիզմում իմունային համակարգը ներկայացված է. ), արյան և ավշային լիմֆոցիտների, ինչպես նաև լիմֆոցիտների և պլազմային բջիջների պոպուլյացիաները, որոնք ներթափանցում են կապի և էպիթելի բոլոր հյուսվածքները: Իմունային համակարգի բոլոր օրգանները գործում են որպես ամբողջություն՝ շնորհիվ նեյրոհումորալ կարգավորիչ մեխանիզմների, ինչպես նաև շրջանառության և ավշային համակարգերի միջոցով բջիջների միգրացիայի և վերաշրջանառության շարունակական գործընթացների: Հիմնական բջիջները, որոնք իրականացնում են մարմնի վերահսկողություն և իմունոլոգիական պաշտպանություն, լիմֆոցիտներն են, ինչպես նաև պլազմային բջիջները և մակրոֆագները:
3. Լիմֆոցիտների երկու հիմնական տեսակ կա՝ B-լիմֆոցիտներ և T-լիմֆոցիտներ: Բ-լիմֆոցիտների ցողունային բջիջները և նախածննդյան բջիջները արտադրվում են ոսկրածուծում: Կաթնասունների մոտ այստեղ տեղի է ունենում B-լիմֆոցիտների տարբերակում, որը բնութագրվում է բջիջներում իմունոգլոբուլինի ընկալիչների ի հայտ գալով։ Ավելին, նման տարբերակված B-լիմֆոցիտները մտնում են ծայրամասային ավշային օրգաններ՝ փայծաղ, ավշային հանգույցներ, մարսողական տրակտի ավշային հանգույցներ: Այս օրգաններում, անտիգենների ազդեցության ներքո, B-լիմֆոցիտները բազմանում են և հետագայում մասնագիտանում են էֆեկտոր բջիջների և հիշողության B-բջիջների ձևավորման հետ:
T-լիմֆոցիտները նույնպես զարգանում են ոսկրածուծի ծագման ցողունային բջիջներից: Վերջիններս արյան հոսքի հետ տեղափոխվում են տիմուս, վերածվում պայթյունների, որոնք բաժանվում ու տարբերվում են երկու ուղղությամբ։ Որոշ պայթյուններ ձևավորում են լիմֆոցիտների պոպուլյացիան հատուկ ընկալիչներով, որոնք ընկալում են օտար անտիգենները: Այս բջիջների տարբերակումը տեղի է ունենում դիֆերենցման ինդուկտորի ազդեցության տակ, որը արտադրվում և արտազատվում է տիմուսի էպիթելային տարրերի կողմից: Ստացված T-լիմֆոցիտները (հակագեն-ռեակտիվ լիմֆոցիտներ) բնակեցնում են ծայրամասային ավշային օրգանների հատուկ T-գոտիներ (տիմուսից կախված): Այնտեղ, անտիգենների ազդեցության տակ, նրանք կարող են փոխակերպվել T-բլաստների, բազմանալ և տարբերակվել փոխպատվաստման մեջ ներգրավված էֆեկտոր բջիջների (T-մարդասպաններ) և հումորալ իմունիտետի (T-օգնականներ և T-suppressors), ինչպես նաև T-ի: հիշողության բջիջներ. T-blast-ի հետնորդների մեկ այլ մասը տարբերվում է սեփական օրգանիզմի անտիգենների համար ընկալիչներ կրող բջիջների ձևավորմամբ: Այս բջիջները ոչնչացվում են:
Այսպիսով, անհրաժեշտ է տարբերակել հակագեն-անկախ և հակագեն-կախյալ տարածումը, B- և T-լիմֆոցիտների տարբերակումը և մասնագիտացումը:
Հյուսվածքային անտիգենների ազդեցությամբ բջջային անձեռնմխելիության ձևավորման դեպքում T-lymphoblasts-ի տարբերակումը հանգեցնում է ցիտոտոքսիկ լիմֆոցիտների (T-killers) և հիշողության T-բջիջների առաջացմանը։ Ցիտոտոքսիկ լիմֆոցիտները ունակ են ոչնչացնել օտար բջիջները (թիրախային բջիջները) կամ հատուկ նյութեր-միջնորդների (լիմֆոկիններ) սեկրեցիայի միջոցով:
Հումորալ իմունիտետի ձևավորման ժամանակ լուծվող և այլ անտիգենների մեծ մասը նույնպես խթանող ազդեցություն ունեն T-լիմֆոցիտների վրա. Միևնույն ժամանակ ձևավորվում են T-օգնականներ, որոնք արտազատում են միջնորդներ (լիմֆոկիններ), որոնք փոխազդում են B-լիմֆոցիտների հետ և առաջացնում նրանց փոխակերպումը B-բլաստների, որոնք մասնագիտացած են պլազմային բջիջների հակամարմիններ արտազատելու գործում: Հակագենով խթանված T-լիմֆոցիտների տարածումը հանգեցնում է նաև բջիջների քանակի ավելացմանը, որոնք վերածվում են ոչ ակտիվ փոքր լիմֆոցիտների, որոնք պահպանում են տեղեկատվություն այս անտիգենի մասին մի քանի տարի և, հետևաբար, կոչվում են հիշողության T-բջիջներ:
T-helper-ը որոշում է B-լիմֆոցիտների մասնագիտացումը հակամարմիններ ձևավորող պլազմային բջիջների ձևավորման ուղղությամբ, որոնք ապահովում են «հումորային իմունիտետ»՝ արտադրելով և արյան մեջ իմունոգոլոբուլիններ արտազատելով: Միևնույն ժամանակ, B-լիմֆոցիտը մակրոֆագից ստանում է հակագենային տեղեկատվություն, որը գրավում է անտիգենը, մշակում և փոխանցում B-լիմֆոցիտին։ B-լիմֆոցիտի մակերեսին ավելի մեծ թվով իմունոգլոբուլինային ընկալիչներ կան (50-150 հազ.)։
Այսպիսով, իմունոլոգիական ռեակցիաներ ապահովելու համար անհրաժեշտ է երեք հիմնական տեսակի բջիջների՝ B-լիմֆոցիտների, մակրոֆագների և T-լիմֆոցիտների գործունեության համագործակցությունը (նկ. 13):
4. Մակրոֆագները կարեւոր դեր են խաղում օրգանիզմի ինչպես բնական, այնպես էլ ձեռքբերովի իմունիտետում։ Մակրոֆագների մասնակցությունը բնական իմունիտետին դրսևորվում է նրանց ֆագոցիտոզ ունակությամբ։ Նրանց դերը ձեռք բերված անձեռնմխելիության մեջ կայանում է նրանում, որ անտիգենը պասիվ փոխանցում է իմունային կոմպետենտ բջիջներին (T- և B-լիմֆոցիտներ)՝ անտիգեններին հատուկ պատասխանի ինդուկցիայի մեջ:
Մակրոֆագների կողմից արտազատվող մշակված հակագենային նյութի մեծ մասը խթանող ազդեցություն ունի T- և B-լիմֆոցիտների կլոնների տարածման և տարբերակման վրա:
Լիմֆյան հանգույցների և փայծաղի B գոտիներում կան մասնագիտացված մակրոֆագներ (դենդրիտիկ բջիջներ), որոնց բազմաթիվ պրոցեսների մակերեսին պահվում են բազմաթիվ անտիգեններ, որոնք մտնում են օրգանիզմ և փոխանցվում B-լիմֆոցիտների համապատասխան կլոններին։ Լիմֆատիկ ֆոլիկուլների T-գոտիներում տեղակայված են միաձուլվող բջիջներ, որոնք ազդում են T-լիմֆոցիտների կլոնների տարբերակման վրա։
Այսպիսով, մակրոֆագները անմիջականորեն ներգրավված են բջիջների (T- և B-լիմֆոցիտներ) համագործակցության մեջ մարմնի իմունային պատասխաններում:
Իմունային համակարգի բջիջների միգրացիայի երկու տեսակ կա՝ դանդաղ և արագ: Առաջինն ավելի բնորոշ է B-լիմֆոցիտներին, երկրորդը՝ T-լիմֆոցիտներին։ Իմունային համակարգի բջիջների միգրացիայի և վերամշակման գործընթացները ապահովում են իմունային հոմեոստազի պահպանումը։
Տես նաև «Կաթնասունների օրգանիզմի պաշտպանական համակարգերի գնահատման մեթոդներ» դասագիրքը (Katsy G.D., Koyuda L.I. - Lugansk. -2003. - p. 42-68):
Կմախքի հյուսվածքներ՝ աճառ և ոսկոր
1. Աճառային հյուսվածքի զարգացումը, կառուցվածքը և տեսակները:
2. Ոսկրային հյուսվածքի զարգացումը, կառուցվածքը և տեսակները.
1. Աճառային հյուսվածքը շարակցական հյուսվածքի մասնագիտացված տեսակ է, որն իրականացնում է օժանդակ ֆունկցիա։ Էմբրիոգենեզում այն զարգանում է մեզենխիմից և ձևավորում է սաղմի կմախքը, որը հետագայում հիմնականում փոխարինվում է ոսկորով: Աճառային հյուսվածքը, բացառությամբ հոդային մակերեսների, ծածկված է խիտ շարակցական հյուսվածքով՝ պերիխոնդրիումով, որը պարունակում է անոթներ, որոնք կերակրում են աճառը և դրա կամբիալ (քոնդրոգեն) բջիջները։
Աճառը բաղկացած է խոնդրոցիտային բջիջներից և միջբջջային նյութից։ Միջբջջային նյութի բնութագրերին համապատասխան՝ աճառի երեք տեսակ կա՝ հիալինային, առաձգական և թելքավոր։
Սաղմի սաղմնային զարգացման գործընթացում մեզենխիմը, ինտենսիվ զարգացող, իրար սերտորեն կից ձևավորում է պրոտոխոնդրալ հյուսվածքային բջիջների կղզիներ։ Նրա բջիջները բնութագրվում են միջուկային-ցիտոպլազմիկ հարաբերակցության բարձր արժեքներով, փոքր խիտ միտոքոնդրիումներով, ազատ ռիբոսոմների առատությամբ, հատիկավոր EPS-ի թույլ զարգացմամբ և այլն: Զարգացման գործընթացում այդ բջիջներից ձևավորվում է առաջնային աճառային (պրախոնդրալ) հյուսվածք: .
Միջբջջային նյութի կուտակման հետ զարգացող աճառի բջիջները մեկուսացվում են առանձին խոռոչներում (լակունաներ) և տարբերվում աճառային հասուն բջիջների՝ քոնդրոցիտների։
Աճառային հյուսվածքի հետագա աճը ապահովվում է խոնդրոցիտների շարունակական բաժանմամբ և դուստր բջիջների միջև միջբջջային նյութի ձևավորմամբ: Վերջինիս ձեւավորումը ժամանակի ընթացքում դանդաղում է։ Դուստր բջիջները, մնալով մեկ բացվածքում, կազմում են բջիջների իզոգեն խմբեր (Isos՝ հավասար, գենեզիս՝ ծագում)։
Քանի որ աճառային հյուսվածքը տարբերվում է, բջիջների վերարտադրության ինտենսիվությունը նվազում է, միջուկները դառնում են պիկտոնիզացված, իսկ միջուկային ապարատը նվազում է:
hyaline աճառ. Հասուն օրգանիզմում հիալինային աճառը կողոսկրերի, կրծքավանդակի մաս է, ծածկում է հոդային մակերեսները և այլն (նկ. 14):
Նրա տարբեր գոտիների աճառային բջիջները՝ քոնդրոցիտները, ունեն իրենց առանձնահատկությունները: Այսպիսով, աճառի ոչ հասուն բջիջները՝ խոնդրոբլաստները, տեղայնացված են անմիջապես պերիխոնդրիումի տակ: Ձվաձեւ են, ցիտոպլազմը հարուստ է ՌՆԹ-ով։ Աճառի ավելի խորը գոտիներում խոնդրոցիտները կլորացվում են՝ ձևավորելով բնորոշ «իզոգեն խմբեր»:
Հիալինային աճառի միջբջջային նյութը պարունակում է ֆիբրիլյար կոլագենային սպիտակուցի չոր քաշի մինչև 70%-ը և ամորֆ նյութի մինչև 30%-ը, որը ներառում է գլիկոզամինոգլիկաններ, պրոտեոգլիկաններ, լիպիդներ և ոչ կոլագենային սպիտակուցներ:
Միջբջջային նյութի մանրաթելերի կողմնորոշումը որոշվում է յուրաքանչյուր աճառին բնորոշ մեխանիկական լարվածության օրինաչափություններով։
Աճառի կոլագենային մանրաթելերը, ի տարբերություն կապի հյուսվածքի այլ տեսակների կոլագենային մանրաթելերի, բարակ են և չեն գերազանցում 10 նմ տրամագիծը:
Աճառային նյութափոխանակությունն ապահովվում է միջբջջային հյուսվածքային հեղուկի շրջանառությամբ, որը կազմում է հյուսվածքների ընդհանուր զանգվածի մինչև 75%-ը։
Էլաստիկ աճառը կազմում է արտաքին ականջի կմախքը, կոկորդի աճառը։ Բացի ամորֆ նյութից և կոլագենի մանրաթելերից, նրա բաղադրությունը ներառում է առաձգական մանրաթելերի խիտ ցանց: Նրա բջիջները նույնական են հիալինային աճառի բջիջներին: Նրանք նաև խմբեր են կազմում և միայնակ պառկում են պերիխոնդրիումի տակ (նկ. 15):
Թելքավոր աճառը տեղայնացված է միջողնաշարային սկավառակների բաղադրության մեջ՝ ջիլը ոսկորներին կցելու տարածքում։ Միջբջջային նյութը պարունակում է կոլագենային մանրաթելերի կոպիտ կապոցներ։ Աճառային բջիջները ձևավորում են իզոգեն խմբեր, որոնք ձգվում են շղթաներով կոլագենի մանրաթելերի կապոցների միջև (նկ. 16):
Աճառային ռեգեներացիան ապահովում է պերիխոնդրիումը, որի բջիջները պահպանում են կամբիալությունը՝ խոնդրոգեն բջիջները։
2. Ոսկրային հյուսվածքը, ինչպես շարակցական հյուսվածքի այլ տեսակներ, զարգանում է մեզենխիմից և բաղկացած է բջիջներից և միջբջջային նյութից։ Այն կատարում է աջակցության, պաշտպանության ֆունկցիա և ակտիվորեն մասնակցում է նյութափոխանակությանը։ Կմախքի սպունգանման ոսկորում տեղայնացված է կարմիր ոսկրածուծը, որտեղ իրականացվում են արյունաստեղծման և օրգանիզմի իմունային պաշտպանության բջիջների տարբերակման գործընթացները։ Ոսկորում կուտակվում են կալցիումի, ֆոսֆորի և այլնի աղեր: Միասին հանքանյութերը կազմում են հյուսվածքի չոր զանգվածի 65-70%-ը:
Ոսկրային հյուսվածքը պարունակում է չորս տարբեր տեսակի բջիջներ՝ օստեոգեն բջիջներ, օստեոբլաստներ, օստեոցիտներ և օստեոկլաստներ:
Օստեոգեն բջիջները օստեոգենեզի գործընթացում մեզենխիմի հատուկ տարբերակման վաղ փուլի բջիջներ են: Նրանք պահպանում են միտոտիկ բաժանման ներուժը: Այս բջիջները տեղայնացված են ոսկրային հյուսվածքի մակերեսին՝ պերիոստեում, էնդոստեում, Հավերսի ջրանցքներում և ոսկրային հյուսվածքի ձևավորման այլ վայրերում։ Բազմանալով՝ նրանք համալրում են օստեոբլաստների պաշարը։
Օստեոբլաստները բջիջներ են, որոնք արտադրում են ոսկրային հյուսվածքի միջբջջային նյութի օրգանական տարրեր՝ կոլագեն, գլիկոզամինոգլիկաններ, սպիտակուցներ և այլն։
Օստեոցիտները գտնվում են միջբջջային նյութի հատուկ խոռոչներում՝ լակունաներում, որոնք փոխկապակցված են բազմաթիվ ոսկրային խողովակներով։
Օստեոկլաստները մեծ, բազմաբնույթ բջիջներ են: Դրանք տեղակայված են ոսկրային հյուսվածքի մակերեսին՝ դրա ռեզորբցման վայրերում։ Բջիջները բևեռացված են: Ներծծվող հյուսվածքի դեմ ուղղված մակերեսը բարակ ճյուղավորման գործընթացների պատճառով ունի ծալքավոր եզրագիծ:
Միջբջջային նյութը բաղկացած է կոլագենային մանրաթելերից և ամորֆ նյութից՝ գլիկոպրոտեիններից, գլիկոզամինոգլիկաններից, սպիտակուցներից և անօրգանական միացություններից։ Մարմնի ողջ կալցիումի 97%-ը կենտրոնացած է ոսկրային հյուսվածքում։
Միջբջջային նյութի կառուցվածքային կազմակերպվածության համաձայն առանձնանում են կոպիտ մանրաթելային ոսկորը և շերտավոր ոսկորը (նկ. 17): Կոպիտ մանրաթելային ոսկորը բնութագրվում է կոլագենի մանրաթելերի կապոցների զգալի տրամագծով և դրանց կողմնորոշման բազմազանությամբ: Բնորոշ է կենդանիների օնտոգենիզացիայի վաղ փուլի ոսկորներին։ Շերտավոր ոսկորում կոլագենի մանրաթելերը կապոցներ չեն կազմում: Զուգահեռ լինելով՝ կազմում են շերտեր՝ ոսկրային թիթեղներ՝ 3-7 մկմ հաստությամբ։ Թիթեղներում կան բջիջների խոռոչներ՝ լակուններ և դրանք միացնող ոսկրային խողովակներ, որոնցում ընկած են օստեոցիտները և դրանց պրոցեսները։ Հյուսվածքային հեղուկը շրջանառվում է լակունների և խողովակների համակարգով, որն ապահովում է նյութափոխանակությունը հյուսվածքում։
Կախված ոսկրային թիթեղների դիրքից՝ առանձնանում են սպունգանման և կոմպակտ ոսկրային հյուսվածքը։ Սպունգային նյութում, մասնավորապես խողովակային ոսկորների էպիֆիզներում, ոսկրային թիթեղների խմբերը տեղակայված են միմյանց նկատմամբ տարբեր անկյուններով։ Քաղցկեղային ոսկրային բջիջները պարունակում են կարմիր ոսկրածուծ:
Կոմպակտ նյութում 4-15 մկմ հաստությամբ ոսկրային թիթեղների խմբերը սերտորեն հարում են միմյանց։ Դիաֆիզում ձևավորվում է երեք շերտ՝ թիթեղների արտաքին ընդհանուր համակարգը, օստեոգեն շերտը և ներքին ընդհանուր համակարգը։
Ծակող խողովակները անցնում են արտաքին ընդհանուր համակարգով պերիոստեումից՝ արյան անոթները և կոլագենային մանրաթելերի կոպիտ կապոցները տեղափոխելով ոսկոր:
Գլանային ոսկորի օստեոգեն շերտում արյունատար անոթներ և նյարդեր պարունակող օստեոնային ուղիները հիմնականում ուղղված են երկայնական: Այս ալիքները շրջապատող խողովակային ոսկրային թիթեղների համակարգը՝ օստեոնները պարունակում են 4-ից 20 թիթեղներ: Օստեոնները միմյանցից սահմանազատվում են հիմնական նյութի ցեմենտի գծով, դրանք ոսկրային հյուսվածքի կառուցվածքային միավորն են (նկ. 18):
Ոսկրային թիթեղների ներքին ընդհանուր համակարգը սահմանակից է ոսկրային շերտի էնդոստեումին և ներկայացված է ջրանցքի մակերեսին զուգահեռ կողմնորոշված թիթեղներով։
Գոյություն ունի օստեոգենեզի երկու տեսակ՝ անմիջապես մեզենխիմից («ուղիղ») և սաղմնային աճառը ոսկրային («անուղղակի») օստեոգենեզով փոխարինելով – թզ. 19.20.
Առաջինը բնորոշ է գանգի և ստորին ծնոտի կոպիտ մանրաթելային ոսկորների զարգացմանը։ Գործընթացը սկսվում է կապի հյուսվածքի և արյան անոթների ինտենսիվ զարգացմամբ: Մեզենխիմային բջիջները, անաստոմոզանալով միմյանց հետ գործընթացներով, կազմում են ցանց։ Միջբջջային նյութի կողմից մակերեսին մի կողմ մղված բջիջները տարբերվում են օստեոբլաստների, որոնք ակտիվորեն մասնակցում են օստեոգենեզին։ Հետագայում առաջնային կոպիտ մանրաթելային ոսկրային հյուսվածքը փոխարինվում է շերտավոր ոսկորով: Աճառային հյուսվածքի տեղում ձևավորվում են կոճղի ոսկորները, վերջույթները և այլն։ Խողովակային ոսկորներում այս գործընթացը սկսվում է դիաֆիզի տարածքում՝ պերիխոնդրիումի տակ գտնվող կոպիտ մանրաթելային ոսկորների խաչաձողերի ցանցի ձևավորմամբ՝ ոսկրային բռունցքով: Աճառը ոսկրային հյուսվածքով փոխարինելու գործընթացը կոչվում է էնդոխոնդրալ ոսկրացում։
Պերիոստեումի կողմից էնդոխոնդրալ ոսկրի զարգացման հետ միաժամանակ տեղի է ունենում պերիխոնդրալ օստեոգենեզի ակտիվ պրոցես՝ ձևավորելով պերիոստեալ ոսկորի խիտ շերտը, որը տարածվում է ամբողջ երկարությամբ մինչև էպիֆիզային աճի թիթեղը։ Պերիոստեալ ոսկորը կմախքի կոմպակտ ոսկրային նյութ է:
Հետագայում ոսկրային էպիֆիզներում հայտնվում են ոսկրացման կենտրոններ։ Ոսկրային հյուսվածքն այստեղ փոխարինում է աճառին։ Վերջինս պահպանվում է միայն հոդային մակերեսին և էպիֆիզային աճի թիթեղում, որը սահմանազատում է էպիֆիզը դիաֆիզից օրգանիզմի աճի ողջ ժամանակահատվածում մինչև կենդանու սեռական հասունացումը։
Պերիոստեումը (periosteum) բաղկացած է երկու շերտից՝ ներքինը պարունակում է կոլագեն և առաձգական մանրաթելեր, օստեոբլաստներ, օստեոկլաստներ և արյունատար անոթներ։ Արտաքին - ձևավորվում է խիտ կապ հյուսվածքով: Այն ուղղակիորեն կապված է մկանների ջլերի հետ։
Endooste - ոսկրածուծի ջրանցքը ծածկող շարակցական հյուսվածքի շերտ: Այն պարունակում է օստեոբլաստներ և կոլագենի մանրաթելերի բարակ կապոցներ, որոնք անցնում են ոսկրածուծի հյուսվածք:
Մկանային հյուսվածքներ
1. Հարթ.
2. Սրտի գծավոր.
3. Կմախքի գծավոր.
4. Մկանային մանրաթելերի զարգացում, աճ և վերականգնում։
1. Մկանային հյուսվածքների առաջատար գործառույթը մարմնի տարածության մեջ տեղաշարժ ապահովելն է որպես ամբողջություն և դրա մասեր: Բոլոր մկանային հյուսվածքները կազմում են մորֆոֆունկցիոնալ խումբ, և կախված կծկվող օրգանելների կառուցվածքից՝ այն բաժանվում է երեք խմբի՝ հարթ, կմախքի գծավոր և սրտի գծավոր մկանային հյուսվածքներ։ Այս հյուսվածքները չունեն սաղմնային զարգացման մեկ աղբյուր։ Դրանք են՝ մեզենխիմը, սեգմենտավորված մեզոդերմայի միոտոմները, սպլանխնոտոմի ներքին օրգանները և այլն։
Մեզենխիմալ ծագման հարթ մկանային հյուսվածք: Հյուսվածքը բաղկացած է միոցիտներից և շարակցական հյուսվածքի բաղադրիչից։ Հարթ միոցիտը 20–500 մկմ երկարությամբ և 5–8 մկմ հաստությամբ միաձույլ բջիջ է։ Ձողաձեւ միջուկը գտնվում է նրա կենտրոնական մասում։ Բջջում կան բազմաթիվ միտոքոնդրիաներ։
Յուրաքանչյուր միոցիտ շրջապատված է նկուղային թաղանթով: Այն ունի բացվածքներ, որոնց շրջանում ձևավորվում են ճեղքաձև հանգույցներ (նեքսուսներ) հարևան միոցիտների միջև՝ ապահովելով միոցիտների ֆունկցիոնալ փոխազդեցությունը հյուսվածքում։ Բազմաթիվ ցանցանման մանրաթելեր հյուսված են նկուղային թաղանթի մեջ։ Մկանային բջիջների շուրջ ցանցային, առաձգական և բարակ կոլագենային մանրաթելերը կազմում են եռաչափ ցանց՝ էնդոմիզիում, որը միավորում է հարևան միոցիտները։
Հարթ մկանային հյուսվածքի ֆիզիոլոգիական ռեգեներացիան սովորաբար դրսևորվում է ֆունկցիոնալ բեռների ավելացման պայմաններում, հիմնականում փոխհատուցվող հիպերտրոֆիայի տեսքով։ Սա առավել հստակ նկատվում է հղիության ընթացքում արգանդի մկանային թաղանթում:
Էպիդերմալ ծագման մկանային հյուսվածքի տարրերը միոէպիթելային բջիջներն են, որոնք զարգանում են էկտոդերմայից: Դրանք տեղակայված են քրտինքի, կաթնագեղձերի, թքագեղձերի և արցունքագեղձերի մեջ՝ միաժամանակ տարբերելով իրենց արտազատվող էպիթելի բջիջների հետ ընդհանուր պրեկուրսորներից։ Կծկվելով՝ բջիջները նպաստում են գեղձի սեկրեցիայի արտազատմանը։
Հարթ մկանները կազմում են մկանային շերտեր բոլոր խոռոչ և խողովակային օրգաններում։
2. Սրտի գծավոր մկանային հյուսվածքի զարգացման աղբյուրները սպլանխնոտոմի վիսցերալ տերեւի սիմետրիկ հատվածներն են։ Նրա բջիջների մեծ մասը տարբերվում է կարդիոմիոցիտների (սրտի միոցիտների), մնացածը՝ էպիկարդիական մեզոթելային բջիջների։ Երկուսն էլ ունեն ընդհանուր նախածննդյան բջիջներ: Հիստոգենեզի ընթացքում տարբերվում են կարդիոմիոցիտների մի քանի տեսակներ՝ կծկվող, հաղորդիչ, անցումային և սեկրետորային։
Կծկվող կարդիոմիոցիտների կառուցվածքը. Բջիջներն ունեն երկարավուն ձև (100-150 մկմ), մոտ գլանաձև։ Նրանց ծայրերը միմյանց հետ կապված են միջանկյալ սկավառակներով։ Վերջիններս կատարում են ոչ միայն մեխանիկական ֆունկցիա, այլև հաղորդիչ, ապահովում են էլեկտրական կապ բջիջների միջև։ Միջուկը օվալաձեւ է, գտնվում է բջջի կենտրոնական մասում։ Այն ունի շատ միտոքոնդրիաներ: Նրանք շղթաներ են կազմում հատուկ օրգանելների՝ միոֆիբրիլների շուրջ։ Վերջիններս կառուցված են ակտինի և միոզինի` կծկվող սպիտակուցների մշտապես առկա պատվիրված թելերից: Դրանք ամրացնելու համար օգտագործվում են հատուկ կառուցվածքներ՝ տելոֆրագմ և մեզոֆրագմ՝ կառուցված այլ սպիտակուցներից։
Երկու Z-գծերի միջև ընկած միոֆիբրիլի հատվածը կոչվում է սարկոմեր: A-bands - անիզոտրոպ, հաստ միկրոթելեր, պարունակում են myosin: I-bands - isotropic, բարակ միկրոթելեր, պարունակում են ակտին; H ժապավենը գտնվում է A-ի միջնամասում (նկ. 21):
Միոցիտների կծկման մեխանիզմի մի քանի տեսություն կա.
1) ցիտոլեմայի միջոցով տարածվող գործողության ներուժի ազդեցությամբ կալցիումի իոններն ազատվում են, մտնում են միոֆիբրիլներ և սկսում կծկվող ակտ, որը ակտինի և միոզինի միկրոթելերի փոխազդեցության արդյունք է. 2) Ներկայումս ամենատարածված տեսությունը սահող թելերի մոդելն է (G. Huxley, 1954): Մենք վերջինիս կողմնակիցներն ենք։
Կարդիոմիոցիտների անցկացման կառուցվածքային առանձնահատկությունները. Բջիջներն ավելի մեծ են, քան աշխատող կարդիոմիոցիտները (երկարությունը մոտ 100 մկմ, իսկ հաստությունը՝ մոտ 50 մկմ): Ցիտոպլազմը պարունակում է ընդհանուր նշանակության բոլոր օրգանելները։ Միոֆիբրիլները քիչ են և գտնվում են բջջի ծայրամասի երկայնքով: Այս կարդիոմիոցիտները միմյանց հետ կապված են մանրաթելերի մեջ ոչ միայն ծայրերում, այլև կողային մակերեսների վրա։ Կարդիոմիոցիտների անցկացման հիմնական գործառույթն այն է, որ նրանք ընկալում են հսկիչ ազդանշանները սրտի ռիթմավարի տարրերից և տեղեկատվություն փոխանցում կծկվող կարդիոմիոցիտներին (նկ. 22):
Վերջնական վիճակում սրտի մկանային հյուսվածքը չի պահպանում ոչ ցողունային բջիջները, ոչ էլ նախածննդյան բջիջները, հետևաբար, եթե կարդիոմիոցիտները մահանում են (սրտի ինֆարկտ), դրանք չեն վերականգնվում:
3. Կմախքի գծավոր մկանային հյուսվածքի տարրերի զարգացման աղբյուրը միոցիտային բջիջներն են։ Նրանցից ոմանք տարբերվում են տեղում, իսկ մյուսները միոտոմներից գաղթում են դեպի մեզենխիմ։ Առաջինները մասնակցում են միոսիմպլաստի ձևավորմանը, երկրորդները տարբերվում են միոսատելիտոցիտների։
Կմախքի մկանային հյուսվածքի հիմնական տարրը մկանային մանրաթելն է, որը ձևավորվում է միոսիմպլաստից և միոսատելիտոցիտներից: Մանրաթելը շրջապատված է սարկոլեմայով։ Քանի որ սիմպլաստը բջիջ չէ, «ցիտոպլազմ» տերմինը չի օգտագործվում, բայց ասում են «սարկոպլազմ» (հունարեն սարկոս՝ միս): Սարկոպլազմայում միջուկների բևեռներում տեղակայված են ընդհանուր նշանակության օրգանելներ։ Հատուկ օրգանոիդները ներկայացված են միոֆիբրիլներով։
Մանրաթելերի կծկման մեխանիզմը նույնն է, ինչ կարդիոմիոցիտներում:
Ներառումները, հիմնականում միոգլոբինը և գլիկոգենը, կարևոր դեր են խաղում մկանային մանրաթելերի գործունեության մեջ։ Գլիկոգենը ծառայում է որպես էներգիայի հիմնական աղբյուր, որն անհրաժեշտ է ինչպես մկանների աշխատանքի, այնպես էլ ամբողջ օրգանիզմի ջերմային հավասարակշռությունը պահպանելու համար։

Բրինձ. 22. Երեք տեսակի կարդիոմիոցիտների ուլտրամիկրոսկոպիկ կառուցվածքը՝ անցկացնող (A), միջանկյալ (B) և աշխատանքային (C) (սխեմա՝ G.S. Katinas)
1 - նկուղային թաղանթ; 2 - բջջային միջուկներ; 3 - միոֆիբրիլներ; 4 - պլազմալեմա; 5 - աշխատանքային կարդիոմիոցիտների միացում (ներդիր սկավառակ); միջանկյալ կարդիոմիոցիտի միացումներ աշխատող և անցկացնող կարդիոմիոցիտների հետ. 6 - անցկացնող կարդիոմիոցիտների միացում; 7 - լայնակի խողովակային համակարգեր (ընդհանուր նշանակության օրգանելները ցուցադրված չեն):
Միոսատելիտոցիտները կից են սիմպլաստի մակերեսին այնպես, որ նրանց պլազմային թաղանթները շփվեն: Արբանյակային բջիջների զգալի քանակությունը կապված է մեկ սիմպլաստի հետ: Յուրաքանչյուր myosatellitocyte միամիջուկային բջիջ է: Միջուկը միոսիմպլաստի միջուկից փոքր է և ավելի կլորացված։ Միտոքոնդրիաները և էնդոպլազմիկ ցանցը հավասարաչափ բաշխված են ցիտոպլազմայում, Գոլջիի համալիրը և բջջային կենտրոնը գտնվում են միջուկի կողքին։ Միոսատելիտոցիտները կմախքի մկանային հյուսվածքի կամբիալ տարրերն են:
Մկանները որպես օրգան. Մկանային մանրաթելերի միջև կան չամրացված շարակցական հյուսվածքի բարակ շերտեր՝ էնդոմիզիում: Դրա ցանցային և կոլագենային մանրաթելերը միահյուսվում են սարկոլեմայի մանրաթելերի հետ, ինչը նպաստում է կծկման ժամանակ ջանքերի միավորմանը։ Մկանային մանրաթելերը խմբավորված են կապոցների մեջ, որոնց միջև կան չամրացված շարակցական հյուսվածքի ավելի հաստ շերտեր՝ պերիմիզիա: Այն նաև պարունակում է առաձգական մանրաթելեր։ Մկանը որպես ամբողջություն շրջապատող շարակցական հյուսվածքը կոչվում է էպիմիզիում:
Անոթայինացում. Մկանային ճյուղ մտնող զարկերակները պերիմիզիայի մեջ: Նրանց կողքին կան բազմաթիվ հյուսվածքային բազոֆիլներ, որոնք կարգավորում են անոթային պատի թափանցելիությունը։ Մազանոթները գտնվում են էնդոմիզիում։ Վեներները և երակները գտնվում են պերիմիզումում՝ զարկերակների և զարկերակների կողքին: Հենց այստեղ են անցնում լիմֆատիկ անոթները։
Իններվացիա. Մկան մտնող նյարդերը պարունակում են ինչպես էֆերենտ, այնպես էլ աֆերենտ մանրաթելեր։ Նյարդային բջջի պրոցեսը, որը բերում է էֆերենտ նյարդային իմպուլսը, ներթափանցում է նկուղային թաղանթով և ճյուղավորվում դրա և սիմպլաստ պլազմոլեմայի միջև՝ մասնակցելով շարժիչի կամ շարժիչային ափսեի ձևավորմանը։ Նյարդային իմպուլսը այստեղ ազատում է միջնորդներ, որոնք առաջացնում են գրգռում, որը տարածվում է սիմպլաստ պլազմալեմայի երկայնքով:
Այսպիսով, յուրաքանչյուր մկանային մանրաթել նյարդայնացվում է ինքնուրույն և շրջապատված է հեմոկափիլյարների ցանցով: Այս համալիրը կազմում է կմախքի մկանների մորֆոֆունկցիոնալ միավորը՝ միոնը; երբեմն մկանային մանրաթելն ինքնին կոչվում է միոն, որը չի համապատասխանում Միջազգային հյուսվածքաբանական անվանացանկին:
4. Բջիջները, որոնցից սաղմոգենեզում առաջանում են գծավոր մկանային մանրաթելեր, կոչվում են միոբլաստներ։ Մի շարք բաժանումներից հետո այս մեկ միջուկային բջիջները, որոնք չեն պարունակում միոֆիբրիլներ, սկսում են միաձուլվել միմյանց հետ՝ ձևավորելով երկարաձգված բազմամիջուկային գլանաձև գոյացություններ՝ միկրոխողովակներ, որոնցում ժամանակին հայտնվում են միոֆիբրիլներ և գծավոր մկանային մանրաթելերին բնորոշ այլ օրգանելներ։ Կաթնասունների մոտ այս մանրաթելերի մեծ մասը ձևավորվում է ծնվելուց առաջ: Հետծննդյան աճի ընթացքում մկանները պետք է դառնան ավելի երկար և հաստ, որպեսզի պահպանեն աճող կմախքի համամասնությունը: Նրանց վերջնական արժեքը կախված է այն աշխատանքից, որը բաժին է ընկնում նրանց: Կյանքի առաջին տարուց հետո մկանների հետագա աճն ամբողջությամբ պայմանավորված է առանձին մանրաթելերի խտացմամբ, այսինքն՝ այն ներկայացնում է հիպերտրոֆիա (հիպեր-ավերջ, ավելի և տրոֆիա-սնուցում), և ոչ թե դրանց քանակի ավելացում, որը կկոչվի հիպերպլազիա: (պլասիսից՝ գոյացում)։
Այսպիսով, գծավոր մկանային մանրաթելերն աճում են հաստությամբ՝ ավելացնելով նրանց մեջ պարունակվող միոֆիբրիլների (և այլ օրգանելների) քանակը։
Արբանյակային բջիջների հետ միաձուլման արդյունքում մկանային մանրաթելերը երկարանում են։ Բացի այդ, հետծննդյան շրջանում հնարավոր է միոֆիբրիլների երկարացում՝ դրանց ծայրերին նոր սարկոմերներ ավելացնելով։
Վերածնում. Արբանյակային բջիջները ոչ միայն ապահովում են գծավոր մկանային մանրաթելերի աճի մեխանիզմներից մեկը, այլև ողջ կյանքի ընթացքում մնում են նոր միոբլաստների պոտենցիալ աղբյուր, որոնց միաձուլումը կարող է հանգեցնել բոլորովին նոր մկանային մանրաթելերի ձևավորմանը: Արբանյակային բջիջները կարող են բաժանվել և առաջացնել միոբլաստներ մկանային վնասվածքից հետո և որոշ դիստրոֆիկ պայմաններում, երբ փորձեր են արվում վերականգնել նոր մանրաթելեր: Այնուամենայնիվ, նույնիսկ ծանր վնասվածքներից հետո մկանային հյուսվածքի աննշան թերությունները լցված են ֆիբրոբլաստների կողմից ձևավորված մանրաթելային հյուսվածքով:
Հարթ մկանների աճ և վերականգնում: Մկանների այլ տեսակների նման, հարթ մկանները արձագանքում են ֆունկցիոնալ պահանջներին փոխհատուցվող հիպերտրոֆիայի միջոցով, բայց սա միակ հնարավոր պատասխանը չէ: Օրինակ՝ հղիության ընթացքում ավելանում է ոչ միայն արգանդի պատի հարթ մկանային բջիջների չափը (հիպերտրոֆիա), այլև դրանց քանակը (հիպերպլազիա)։
Կենդանիների մոտ հղիության ընթացքում կամ հորմոնների ընդունումից հետո, արգանդի մկանային բջիջներում հաճախ կարելի է տեսնել միտոտիկ թվեր. հետևաբար, ընդհանուր առմամբ ընդունված է, որ հարթ մկանային բջիջները պահպանում են միտոտիկ բաժանման ունակությունը:
նյարդային հյուսվածք
1. Հյուսվածքների զարգացում.
2. Նյարդային բջիջների դասակարգում.
3. Նեյրոգլիա, նրա բազմազանությունը:
4. Սինապսներ, մանրաթելեր, նյարդային վերջավորություններ:
1. Նյարդային հյուսվածք՝ մասնագիտացված հյուսվածք, որը կազմում է օրգանիզմի հիմնական ինտեգրացիոն համակարգը՝ նյարդային համակարգը։ Հիմնական գործառույթը հաղորդունակությունն է:
Նյարդային հյուսվածքը բաղկացած է նյարդային բջիջներից՝ նեյրոններից, որոնք կատարում են նյարդային գրգռման և նյարդային իմպուլսի անցկացման գործառույթը, և նեյրոգլիա, որն ապահովում է օժանդակ, տրոֆիկ և պաշտպանիչ գործառույթներ:
Նյարդային հյուսվածքը զարգանում է էկտոդերմի մեջքային խտացումից՝ նյարդային թիթեղից, որը զարգացման գործընթացում տարբերվում է նյարդային խողովակի, նյարդային գագաթների (գլանիկների) և նյարդային պլակոդների։
Սաղմի առաջացման հետագա ժամանակաշրջաններում ուղեղը և ողնուղեղը ձևավորվում են նյարդային խողովակից: Նյարդային գագաթը ձևավորում է զգայական գանգլիաներ, սիմպաթիկ նյարդային համակարգի գանգլիաներ, մաշկի մելանոցիտներ և այլն: Նյարդային պլակոդները ներգրավված են հոտի, լսողության և զգայական գանգլիաների ձևավորման մեջ:
Նյարդային խողովակը բաղկացած է պրիզմատիկ բջիջների մեկ շերտից: Վերջիններս, բազմանալով, կազմում են երեք շերտ՝ ներքին՝ էպենդիմալ, միջին՝ թիկնոց և արտաքին՝ եզրային շղարշ։
Հետագայում ներքին շերտի բջիջները արտադրում են ողնուղեղի կենտրոնական ջրանցքը ծածկող էպենդիմալ բջիջներ։ Թիկնոցի շերտի բջիջները տարբերվում են նեյրոբլաստների, որոնք հետագայում վերածվում են նեյրոնների և սպունգիոբլաստների՝ առաջացնելով տարբեր տեսակի նեյրոգլիաներ (աստրոցիտներ, օլիգոդենդրոցիտներ)։
2. Նյարդային համակարգի տարբեր մասերի նյարդային բջիջները (նեյրոցիտներ, նեյրոններ) բնութագրվում են տարբեր ձևերով, չափերով և ֆունկցիոնալ նշանակությամբ։ Նյարդային բջիջները, ըստ ֆունկցիայի, բաժանվում են ընկալիչի (աֆերենտ), ասոցիատիվ և էֆեկտորային (էֆերենտ):
Նյարդային բջիջների ձևերի լայն տեսականիով, ընդհանուր ձևաբանական առանձնահատկությունն այն գործընթացների առկայությունն է, որոնք ապահովում են դրանց կապը որպես ռեֆլեքսային աղեղների մաս: Գործընթացների երկարությունը տարբեր է և տատանվում է մի քանի միկրոնից մինչև 1-1,5 մ:
Նյարդային բջիջների պրոցեսները ըստ ֆունկցիոնալ նշանակության բաժանվում են երկու տեսակի. Ոմանք ստանում են նյարդային գրգռում և այն տեղափոխում նեյրոնի պերիկարիոն: Դրանք կոչվում են դենդրիտներ։ Մեկ այլ տեսակի պրոցեսներ իմպուլս են փոխանցում բջջային մարմնից և փոխանցում այն մեկ այլ նեյրոցիտ կամ աքսոն (աքսոս - առանցք) կամ նեյրիտ: Բոլոր նյարդային բջիջներն ունեն միայն մեկ նեյրիտ:
Ըստ պրոցեսների քանակի՝ նյարդային բջիջները բաժանվում են միաբևեռ՝ մեկ պրոցեսով՝ երկբևեռ և բազմաբևեռ (նկ. 23)։
Նյարդային բջիջների միջուկները մեծ են, կլորացված կամ թեթևակի օվալաձև, գտնվում են պերիկարիոնի կենտրոնում։
Բջիջների ցիտոպլազման բնութագրվում է տարբեր օրգանելների, նեյրոֆիբրիլների և քրոմատոֆիլ նյութերի առատությամբ։ Բջջի մակերեսը ծածկված է պլազմալեմայով, որը բնութագրվում է գրգռվածությամբ և գրգռում անցկացնելու ունակությամբ։

Բրինձ. 23. Նյարդային բջիջների տեսակները (սխեմա ըստ T.N. Radostina, L.S. Rumyantseva)
A - միաբևեռ նեյրոն; B - pseudounipolar neuron; B - երկբևեռ նեյրոն; D - բազմաբևեռ նեյրոն:
Նեյրոֆիբրիլները մանրաթելերի հավաքածու են, ցիտոպլազմայի կառուցվածքներ, որոնք կազմում են պերիկարիոնում խիտ պլեքսուս:
Քրոմատոֆիլ (բազոֆիլ) նյութը հայտնաբերվում է նեֆրոցիտների պերիկարիոնում և նրանց դենդրիտներում, սակայն բացակայում է աքսոններում։
Էպենդիմոցիտները գծում են կենտրոնական նյարդային համակարգի խոռոչները՝ ուղեղի փորոքները և ողնաշարի ջրանցքը: Նյարդային խողովակի խոռոչի դեմ ուղղված բջիջները պարունակում են թարթիչներ: Նրանց հակառակ բևեռները անցնում են երկար գործընթացների, որոնք աջակցում են նյարդային խողովակի հյուսվածքների ողնաշարին: Էպենդիմոցիտները ներգրավված են արտազատման ֆունկցիայի մեջ՝ ազատելով տարբեր ակտիվ նյութեր արյան մեջ։
Աստրոցիտները կամ պրոտոպլազմային են (կարճ ճառագայթով) կամ մանրաթելային (երկար ճառագայթով): Առաջինները տեղայնացված են Կենտրոնական նյարդային համակարգի (կենտրոնական նյարդային համակարգ) գորշ հարցում: Նրանք ներգրավված են նյարդային հյուսվածքի նյութափոխանակության մեջ և կատարում են սահմանազատող ֆունկցիա։
ԿՆՀ-ի սպիտակ նյութին բնորոշ են թելքավոր աստրոցիտները։ Նրանք կազմում են կենտրոնական նյարդային համակարգի օժանդակ ապարատը:
Օլիգոդենդրոցիտները CNS և PNS (ծայրամասային նյարդային համակարգ) բջիջների մեծ խումբ են: Նրանք շրջապատում են նեյրոնների մարմինները, մտնում են նյարդաթելերի և նյարդային վերջավորությունների պատյանների մեջ և մասնակցում դրանց նյութափոխանակությանը։
Microglia (glial macrophages) մակրոֆագների մասնագիտացված համակարգ է, որն իրականացնում է պաշտպանիչ գործառույթ: Զարգանում են մեզենխիմից, ընդունակ են ամեոբային շարժման։ Դրանք բնորոշ են ԿՆՀ-ի սպիտակ և մոխրագույն նյութին։
4. Նյարդային բջիջների պրոցեսները դրանք ծածկող նեյրոգլիայի հետ միասին կազմում են նյարդաթելեր։ Դրանցում տեղակայված նյարդային բջիջների պրոցեսները կոչվում են առանցքային բալոններ, իսկ դրանք ծածկող օլիգոդենդրոգլիա բջիջները՝ նեյրոլեմմոցիտներ (Շվանի բջիջներ)։
Տարբերակել միելինացված և ոչ միելինացված նյարդաթելերը:
Ինքնավար նյարդային համակարգին բնորոշ են ոչ միելինացված (ոչ միելինացված) նյարդաթելերը։ Լեմմոցիտները սերտորեն տեղավորվում են միմյանց հետ, ձևավորելով շարունակական թելեր: Մանրաթելը պարունակում է մի քանի առանցքային բալոններ, այսինքն, տարբեր նյարդային բջիջների գործընթացներ: Պլազմալեմման ձևավորում է խոր ծալքեր, որոնք կազմում են կրկնակի թաղանթ՝ մեսաքսոն, որի վրա կախված է առանցքային գլան։ Լույսի մանրադիտակով այս կառույցները չեն հայտնաբերվում, ինչը ստեղծում է առանցքային բալոնների ընկղմման տպավորություն անմիջապես գլիալ բջիջների ցիտոպլազմայի մեջ:
Myelinated (pulp) նյարդաթելեր. Նրանց տրամագիծը տատանվում է 1-ից 20 մկմ: Դրանք պարունակում են մեկ առանցքային գլան՝ նյարդային բջջի դենդրիտ կամ նեյրիտ՝ ծածկված լեմմոցիտներով ձևավորված պատյանով։ Մանրաթելային թաղանթում առանձնանում են երկու շերտ՝ ներքինը միելին է, ավելի հաստ, իսկ արտաքինը՝ բարակ, պարունակում է լեմոցիտների ցիտոպլազմա և միջուկներ։
Երկու լեմմոցիտների սահմանին միելինային մանրաթելերի պատյանը բարակվում է, ձևավորվում է մանրաթելի նեղացում՝ հանգույցային ընդհատում (Ranvier interception): Նյարդային մանրաթելի երկու հատվածների միջև ընկած հատվածը կոչվում է միջնոդալ հատված: Նրա կեղևը համապատասխանում է մեկ լեմմոցիտի:
Նյարդային վերջավորությունները տարբեր են իրենց ֆունկցիոնալ նշանակությամբ։ Գոյություն ունեն նյարդային վերջավորությունների երեք տեսակ՝ էֆեկտոր, ընկալիչ և վերջավոր սարքեր։
Էֆեկտորային նյարդերի վերջավորություններ - դրանք ներառում են գծավոր և հարթ մկանների շարժիչային նյարդերի վերջավորությունները և գեղձային օրգանների արտազատիչ վերջավորությունները:
Կմախքի գծավոր մկանների շարժիչային նյարդերի վերջավորություններ - շարժիչային սալիկներ - նյարդային և մկանային հյուսվածքների փոխկապակցված կառուցվածքների համալիր:
Զգայական նյարդերի վերջավորությունները (ընկալիչները) զգայուն նեյրոնների դենդրիտների մասնագիտացված տերմինալ կազմավորումներ են։ Գոյություն ունեն ընկալիչների երկու մեծ խմբեր՝ էքստերորեսեպտորներ և միջընկալիչներ։ Զգայական վերջավորությունները բաժանվում են մեխանոռեցեպտորների, քիմոընկալիչների, ջերմընկալիչների և այլն, բաժանվում են ազատ և ոչ ազատ նյարդային վերջավորությունների։ Վերջիններս ծածկված են շարակցական հյուսվածքի պարկուճով և կոչվում են պարկուճ։ Այս խումբը ներառում է շերտավոր մարմիններ (Վատեր-Պաչինի մարմիններ), շոշափելի մարմիններ (Մայսների մարմիններ) և այլն։
Շերտավոր մարմինները բնորոշ են մաշկի խորքային շերտերին և ներքին օրգաններին։ Շոշափելի մարմինները ձևավորվում են նաև գլիալ բջիջներով։
Սինապսները երկու նեյրոնների մասնագիտացված շփում են, որոնք ապահովում են նյարդային գրգռման միակողմանի փոխանցում: Ձևաբանորեն սինապսում առանձնանում են նախասինապսային և հետսինապսային բևեռները, որոնց միջև կա անջրպետ։ Կան սինապսներ՝ քիմիական և էլեկտրական փոխանցման միջոցով։
Ըստ շփման վայրի՝ առանձնացնում են սինապսները՝ աքսոսոմատիկ, աքսոդենդրիալ և աքսոաքսոնալ։
Սինապսի նախասինապսային բևեռը բնութագրվում է միջնորդ (ացետիլխոլին կամ նորէպինեֆրին) պարունակող սինապտիկ վեզիկուլների առկայությամբ:
Նյարդային համակարգը ներկայացված է զգայական և շարժիչ բջիջներով, որոնք միավորված են միջնեյրոնային սինապսներով՝ ֆունկցիոնալ ակտիվ կազմավորումների՝ ռեֆլեքսային աղեղների։ Պարզ ռեֆլեքսային աղեղը բաղկացած է երկու նեյրոնից՝ զգայական և շարժիչ:
Բարձր ողնաշարավորների ռեֆլեքսային աղեղները դեռևս պարունակում են զգալի թվով ասոցիատիվ նեյրոններ, որոնք տեղակայված են զգայական և շարժիչ նեյրոնների միջև։
Նյարդը մանրաթելերի փաթեթ է, որը շրջապատված է պերինևրիումի խիտ պատյանով։ Փոքր նյարդերը բաղկացած են միայն մեկ կապոցից, որը շրջապատված է էնդոնևրիումով: Փաթեթում նյարդային մանրաթելերի քանակը և տրամագիծը շատ փոփոխական են: Որոշ նյարդերի հեռավոր հատվածներում կան ավելի շատ մանրաթելեր, քան ավելի մոտակա հատվածներում: Դա պայմանավորված է մանրաթելերի ճյուղավորմամբ:
Արյան մատակարարում նյարդերին. Նյարդերը առատորեն մատակարարվում են բազմաթիվ անաստոմոզներ ձևավորող անոթներով։ Տարբերում են էպինևրալ, միջֆասկուլյար, պերինևրալ և ներֆասիկուլյար զարկերակներ և զարկերակներ։ Endoneurium-ը պարունակում է մազանոթների ցանց։
գրականություն
1. Ալեքսանդրովսկայա Օ.Վ., Ռադոստինա Տ.Ն., Կոզլով Ն.Ա. Բջջաբանություն, հյուսվածքաբանություն և սաղմնաբանություն.-M: Agropromizdat, 1987.- 448 p.
2. Աֆանասիև Յու.Ի., Յուրինա Ն.Ա. Histology.- M: Բժշկություն, 1991.- 744 p.
3. Վրակին Վ.Ֆ., Սիդորովա Մ.Վ. Գյուղատնտեսական կենդանիների մորֆոլոգիա. - M: Agropromizdat, 1991.- 528 p.
4. Գլագոլև Պ.Ա., Իպոլիտովա Վ.Ի. Գյուղատնտեսական կենդանիների անատոմիա՝ հյուսվածաբանության և սաղմնաբանության հիմունքներով:- M: Kolos, 1977.- 480 p.
5. Ham A., Cormac D. Histology. -M: Mir, 1982.-T 1-5.
6. Սերավին Լ.Ն. Էուկարիոտիկ բջջի ծագումը //Citology.-1986/-T. 28.-Թիվ 6-8.
7. Սերավին Լ.Ն. Բջջային տեսության զարգացման հիմնական փուլերը և բջջի տեղը կենդանի համակարգերում //Citology.-1991.-V.33.-№ 12/-C. 3-27։
Հյուսվածքը բջիջների և ոչ բջջային կառուցվածքների համակարգ է, որն առաջացել է էվոլյուցիայի գործընթացում՝ միավորված ընդհանուր կառուցվածքով և գործառույթներով (ցանկալի է իմանալ սահմանումը և հասկանալ իմաստը. 1) հյուսվածքն առաջացել է էվոլյուցիայի գործընթաց, 2) դա բջիջների և ոչ բջջային կառուցվածքների համակարգ է, 3) կա ընդհանուր կառուցվածք, 4) բջիջների և ոչ բջջային կառուցվածքների համակարգ, որոնք տվյալ հյուսվածքի մաս են կազմում և ունեն ընդհանուր գործառույթներ):Կառուցվածքային և ֆունկցիոնալ տարրերհյուսվածքները բաժանվում են. հյուսվածքաբանական տարրեր բջջային (1)Եվ ոչ բջջային տիպ (2). Հյուսվածքների կառուցվածքային և ֆունկցիոնալ տարրեր մարդու մարմինըկարելի է համեմատել տեքստիլ գործվածքներից կազմված տարբեր թելերի հետ։
«Հիալինային աճառ» հյուսվածաբանական պատրաստուկ՝ 1-քոնդրոցիտային բջիջներ, 2-միջբջջային նյութ (ոչ բջջային տիպի հյուսվածաբանական տարր)
1. Բջջի տիպի հյուսվածքաբանական տարրերսովորաբար կենդանի կառուցվածքներ են՝ իրենց սեփական նյութափոխանակությամբ, սահմանափակված պլազմային թաղանթով, և հանդիսանում են բջիջներ և դրանց ածանցյալներ, որոնք առաջանում են մասնագիտացման արդյունքում: Դրանք ներառում են.
Ա) Բջիջներ- հյուսվածքների հիմնական տարրերը, որոնք որոշում են դրանց հիմնական հատկությունները.
բ) Հետբջջային կառուցվածքներորոնցում կորչում են բջիջների համար ամենակարևոր նշանները (միջուկ, օրգանելներ), օրինակ՝ էրիթրոցիտները, էպիդերմիսի եղջյուրավոր թեփուկները, ինչպես նաև թրոմբոցիտները, որոնք բջիջների մասեր են.
V) Սիմպլաստներ- առանձին բջիջների միաձուլման արդյունքում ձևավորված կառույցներ՝ բազմաթիվ միջուկներով և ընդհանուր պլազմային թաղանթով մեկ ցիտոպլազմային զանգվածի մեջ, օրինակ՝ կմախքի մկանային հյուսվածքի մանրաթել, օստեոկլաստ.
է) սինցիտիա- կառուցվածքներ, որոնք բաղկացած են բջիջներից, որոնք միավորված են մեկ ցանցում ցիտոպլազմային կամուրջներով՝ թերի տարանջատման պատճառով, օրինակ՝ սպերմատոգեն բջիջները վերարտադրության, աճի և հասունացման փուլերում։
2. Ոչ բջջային տիպի հյուսվածքաբանական տարրերներկայացված են նյութերով և կառուցվածքներով, որոնք արտադրվում են բջիջների կողմից և ազատվում պլազմալեմից դուրս՝ միավորված ընդհանուր անվան տակ։ «միջբջջային նյութ» (հյուսվածքային մատրիցա): միջբջջային նյութսովորաբար ներառում է հետևյալ սորտերը.
Ա) Ամորֆ (հիմնական) նյութ – ներկայացված է օրգանական (գլիկոպրոտեիններ, գլիկոզամինոգլիկաններ, պրոտեոգլիկաններ) և անօրգանական (աղեր) նյութերի անկառույց կուտակմամբ, որոնք տեղակայված են հյուսվածքային բջիջների միջև հեղուկ, գելանման կամ պինդ, երբեմն բյուրեղացված վիճակում (ոսկրային հյուսվածքի հիմնական նյութը).
բ) մանրաթելեր – բաղկացած են ֆիբրիլային սպիտակուցներից (էլաստին, տարբեր տեսակի կոլագեն), որոնք հաճախ ամորֆ նյութում ձևավորում են տարբեր հաստության կապոցներ։ Դրանց թվում առանձնանում են. 1) կոլագեն, 2) ռետիկուլյար և 3) առաձգական մանրաթելեր. Fibrillar սպիտակուցները ներգրավված են նաև բջջային պարկուճների (աճառ, ոսկորներ) և նկուղային թաղանթների (էպիթելի) ձևավորման մեջ:
 Լուսանկարում ներկայացված է «Չամրացված թելքավոր շարակցական հյուսվածք» հյուսվածաբանական պատրաստուկը՝ հստակ տեսանելի են բջիջները, որոնց միջև առկա է միջբջջային նյութը (մանրաթելեր՝ շերտեր ամորֆ նյութթեթև տարածքներ բջիջների միջև):
Լուսանկարում ներկայացված է «Չամրացված թելքավոր շարակցական հյուսվածք» հյուսվածաբանական պատրաստուկը՝ հստակ տեսանելի են բջիջները, որոնց միջև առկա է միջբջջային նյութը (մանրաթելեր՝ շերտեր ամորֆ նյութթեթև տարածքներ բջիջների միջև):
2. Գործվածքների դասակարգում. Համաձայն մորֆոֆունկցիոնալ դասակարգումառանձնանում են հյուսվածքներ՝ 1) էպիթելային հյուսվածքներ, 2) ներքին միջավայրի հյուսվածքներ՝ շարակցական և արյունաստեղծ, 3) մկանային և 4) նյարդային հյուսվածք։
3. Հյուսվածքների զարգացում. Տարբեր զարգացման տեսությունգործվածքներ ըստ Ն.Գ. Խլոպինը ենթադրում է, որ հյուսվածքները առաջացել են դիվերգենցիայի արդյունքում՝ նշանների տարաձայնություն՝ կապված կառուցվածքային բաղադրիչների հարմարեցման նոր պայմանների հետ։ Զուգահեռ շարքերի տեսությունըստ Ա.Ա. Զավարզինը նկարագրում է հյուսվածքների էվոլյուցիայի պատճառները, որոնց համաձայն նմանատիպ գործառույթներ կատարող հյուսվածքները ունեն նմանատիպ կառուցվածք։ Ֆիլոգենեզի ընթացքում կենդանական աշխարհի տարբեր էվոլյուցիոն ճյուղերում զուգահեռաբար առաջացել են նույնական հյուսվածքներ, այսինքն. բնօրինակ հյուսվածքների բոլորովին տարբեր ֆիլոգենետիկ տիպերը, ընկնելով արտաքին կամ ներքին միջավայրի գոյության համանման պայմանների մեջ, տվել են նմանատիպ մորֆոֆունկցիոնալ տիպի հյուսվածքներ։ Այս տեսակները ֆիլոգենիայում առաջանում են միմյանցից անկախ, այսինքն. զուգահեռաբար, էվոլյուցիայի նույն հանգամանքներում կենդանիների բացարձակապես տարբեր խմբերում: Այս երկու փոխլրացնող տեսությունները միավորվում են մեկում Հյուսվածքների էվոլյուցիոն հայեցակարգ(Ա.Ա. Բրաուն և Պ.Պ. Միխայլով), ըստ որի ֆիլոգենետիկ ծառի տարբեր ճյուղերում միանման հյուսվածքային կառուցվածքներ առաջացել են զուգահեռաբար դիվերգենտ զարգացման ընթացքում։
Ինչպե՞ս կարող է նման բազմազան կառուցվածքներ ձևավորվել մեկ բջջից՝ զիգոտից: Դրա համար պատասխանատու են այնպիսի գործընթացներ, ինչպիսիք են ՈՐՈՇՈՒՄԸ, ՊԱՐՏԱՎՈՐՈՒՄԸ, ՏԱՐԲԵՐԱԿՈՒՄԸ: Փորձենք հասկանալ այս տերմինները։
վճռականություն- սա մի գործընթաց է, որը որոշում է սաղմնային ռուդիմենտներից բջիջների, հյուսվածքների զարգացման ուղղությունը: Որոշման ընթացքում բջիջները հնարավորություն են ստանում զարգանալ որոշակի ուղղությամբ։ Արդեն զարգացման վաղ փուլերում, երբ ջախջախումը տեղի է ունենում, հայտնվում են երկու տեսակի բլաստոմերներ՝ բաց և մութ։ Թեթև բլաստոմերներից, օրինակ, կարդիոմիոցիտները և նեյրոնները հետագայում չեն կարող ձևավորվել, քանի որ դրանք որոշվում են և դրանց զարգացման ուղղությունը քորիոնային էպիթելիումն է: Այս բջիջները զարգանալու շատ սահմանափակ հնարավորություններ (պոտենցիա) ունեն։
Օրգանիզմի զարգացման ծրագրին համահունչ քայլ առ քայլ կոչվում է որոշման պատճառով զարգացման հնարավոր ուղիների սահմանափակում. պարտավորվելով . Օրինակ, եթե երկշերտ սաղմի առաջնային էկտոդերմի բջիջները դեռ կարող են զարգացնել երիկամային պարենխիմայի բջիջները, ապա երկրորդական էկտոդերմայից եռաշերտ սաղմի (էկտո-, մեզո- և էնդոդերմա) հետագա զարգացմամբ և ձևավորմամբ, միայն նյարդային հյուսվածք, մաշկի էպիդերմիս և որոշ այլ բաներ:
Մարմնի բջիջների և հյուսվածքների որոշումը, որպես կանոն, անշրջելի է. մեզոդերմային բջիջները, որոնք առաջնային շերտից դուրս են եկել երիկամային պարենխիման ձևավորելու համար, չեն կարողանա նորից վերածվել առաջնային էկտոդերմային բջիջների:
Տարբերակումուղղված է բազմաբջջային օրգանիզմում մի քանի կառուցվածքային և ֆունկցիոնալ բջիջների տիպերի ստեղծմանը: Մարդկանց մոտ այդպիսի բջիջների ավելի քան 120 տեսակ կա։Դիֆերենցման ընթացքում տեղի է ունենում հյուսվածքային բջիջների մասնագիտացման մորֆոլոգիական և ֆունկցիոնալ նշանների աստիճանական ձևավորում (բջիջների տեսակների ձևավորում)։
Դիֆերոննույն տիպի բջիջների հիստոգենետիկ շարքն է տարբերակման տարբեր փուլերում: Ինչպես ավտոբուսում գտնվող մարդիկ՝ երեխաներ, երիտասարդներ, մեծահասակներ, տարեցներ: Եթե ավտոբուսով կատու և ձագեր են տեղափոխում, ապա կարելի է ասել, որ ավտոբուսում կա «երկու դիֆերոն»՝ մարդիկ և կատուներ:
Որպես Differon-ի մաս՝ ըստ տարբերակման աստիճանի առանձնանում են հետևյալ բջիջների պոպուլյացիաները. Ցողունային բջիջները- տվյալ հյուսվածքի ամենաքիչ տարբերակված բջիջները, որոնք կարող են բաժանվել և լինել նրա մյուս բջիջների զարգացման աղբյուր. բ) կիսա-ցողունային բջիջներ- պրեկուրսորները պարտավորվածության պատճառով տարբեր տեսակի բջիջներ ձևավորելու իրենց ունակության սահմանափակումներ ունեն, բայց կարող են ակտիվ վերարտադրվել. V) բջիջները պայթյուններ ենորոնք մտել են տարբերակման մեջ, բայց պահպանում են բաժանելու ունակությունը. է) հասունացող բջիջները- լրացնել տարբերակումը; ե) հասունացած(տարբերակված) բջիջները, որոնք լրացնում են հիստոգենետիկ շարքը, նրանց բաժանվելու ունակությունը, որպես կանոն, անհետանում է, նրանք ակտիվորեն գործում են հյուսվածքում. ե) հին բջիջներ- ավարտված ակտիվ գործողություն:
Բջիջների մասնագիտացման մակարդակը դիֆերոնային պոպուլյացիաներում աճում է ցողունային բջիջներից մինչև հասուն բջիջներ: Այս դեպքում տեղի են ունենում ֆերմենտների, բջիջների օրգանելների բաղադրության և ակտիվության փոփոխություններ։ Դիֆերոնի հիստոգենետիկ շարքը բնութագրվում է տարբերակման անշրջելիության սկզբունքը, այսինքն. նորմալ պայմաններում ավելի տարբերակված վիճակից ավելի քիչ տարբերակված վիճակի անցումն անհնար է։ Դիֆերոնի այս հատկությունը հաճախ խախտվում է, երբ պաթոլոգիական պայմաններ(չարորակ ուռուցքներ):
Մկանային մանրաթելի ձևավորմամբ կառուցվածքների տարբերակման օրինակ (զարգացման հաջորդական փուլեր):
Զիգոտ - բլաստոցիստ - ներքին բջիջների զանգված (էմբրիոբլաստ) - էպիբլաստ - մեզոդերմա - չհատված մեսոդերմա- սոմիտ - somite myotome բջիջները- միտոտիկ միոբլաստներ - պոստմիտոտիկ միոբլաստներ - մկանային խողովակ - մկանային մանրաթել:
Վերոնշյալ սխեմայում փուլից փուլ սահմանափակվում է տարբերակման հնարավոր ուղղությունների թիվը։ Բջիջներ չհատված մեսոդերմաունեն տարբեր ուղղություններով տարբերվելու կարողություն (պոտենցիա) և ձևավորելու միոգեն, քոնդրոգեն, օստեոգեն և այլ ուղղություններ։ Somite myotome բջիջներըորոշված են զարգանալ միայն մեկ ուղղությամբ, այն է՝ միոգեն բջիջների տիպի ձևավորում (կմախքի տիպի գծավոր մկան):
Բջջային պոպուլյացիաներօրգանիզմի կամ հյուսվածքի բջիջների հավաքածու է, որոնք ինչ-որ կերպ նման են միմյանց։ Ըստ բջիջների բաժանման միջոցով ինքնավերականգնման ունակության, առանձնանում են բջիջների պոպուլյացիաների 4 կատեգորիաներ (ըստ Լեբլոնի).
- Սաղմնային(արագ բաժանվող բջիջների պոպուլյացիա) - բնակչության բոլոր բջիջները ակտիվորեն բաժանվում են, մասնագիտացված տարրերը բացակայում են:
- կայունԲջջային պոպուլյացիա՝ երկարակյաց, ակտիվ գործող բջիջներ, որոնք ծայրահեղ մասնագիտացման պատճառով կորցրել են բաժանվելու ունակությունը։ Օրինակ՝ նեյրոնները, կարդիոմիոցիտները։
- Աճում(անկայուն) բջիջների պոպուլյացիա - մասնագիտացված բջիջներ, որոնք ի վիճակի են բաժանվել որոշակի պայմաններում: Օրինակ, երիկամի, լյարդի էպիթելը:
- Բնակչության բարելավումբաղկացած է անընդհատ և արագ բաժանվող բջիջներից, ինչպես նաև այդ բջիջների մասնագիտացված գործող ժառանգներից, որոնց կյանքի տևողությունը սահմանափակ է։ Օրինակ՝ աղիքային էպիթելը, արյունաստեղծ բջիջները։
Բջջային պոպուլյացիաների հատուկ տեսակ են կլոն- նույնական բջիջների խումբ, որը ստացվում է մեկ նախնիների նախահայր բջիջից: հայեցակարգը կլոնորպես բջջային պոպուլյացիան հաճախ օգտագործվում է իմունոլոգիայում, օրինակ՝ T-լիմֆոցիտների կլոնը։
4. Հյուսվածքների վերածնում- գործընթաց, որն ապահովում է դրա նորացումը նորմալ կյանքի ընթացքում (ֆիզիոլոգիական վերածնում) կամ վնասից հետո վերականգնում (վերականգնողական վերականգնում):
կամբիալ տարրեր - սրանք ցողունային, կիսա-ցողունային նախաբջիջների պոպուլյացիաներ են, ինչպես նաև տվյալ հյուսվածքի պայթուցիկ բջիջներ, որոնց բաժանումը պահպանում է իր բջիջների անհրաժեշտ քանակը և լրացնում հասուն տարրերի պոպուլյացիայի անկումը: Այն հյուսվածքներում, որտեղ բջիջների նորացումը տեղի չի ունենում բջիջների բաժանման միջոցով, կամբիումը բացակայում է։ Ըստ կամբիալ հյուսվածքի տարրերի բաշխման՝ առանձնանում են կամբիումի մի քանի տեսակներ.
- տեղայնացված կամբիում– նրա տարրերը կենտրոնացած են հյուսվածքի որոշակի հատվածներում, օրինակ՝ շերտավորված էպիթելում, կամբիումը տեղայնացված է բազալային շերտում.
- ցրված կամբիում– դրա տարրերը ցրված են հյուսվածքի մեջ, օրինակ՝ հարթ մկանային հյուսվածքում, կամբիալ տարրերը ցրված են տարբերակված միոցիտների միջև.
- Բացահայտված կամբիում- նրա տարրերը գտնվում են հյուսվածքից դուրս և, քանի որ դրանք տարբերվում են, ներառված են հյուսվածքի կազմի մեջ, օրինակ՝ արյունը պարունակում է միայն տարբերակված տարրեր, կամբիումի տարրերը տեղակայված են արյունաստեղծ օրգաններում։
Հյուսվածքների վերականգնման հնարավորությունը որոշվում է նրա բջիջների՝ բաժանվելու և տարբերվելու ունակությամբ կամ ներբջջային վերածնման մակարդակով։ Հյուսվածքները, որոնք ունեն կամբիալ տարրեր կամ նորանում կամ աճում են բջիջների պոպուլյացիաները, լավ վերականգնվում են: Վերականգնման ընթացքում յուրաքանչյուր հյուսվածքի բջիջների բաժանման (տարածման) ակտիվությունը վերահսկվում է աճի գործոններով, հորմոններով, ցիտոկիններով, կալոններով, ինչպես նաև ֆունկցիոնալ բեռների բնույթով:
Բացի բջիջների բաժանման միջոցով հյուսվածքների և բջիջների վերածնումից, կա ներբջջային վերածնում- բջջի կառուցվածքային բաղադրիչների շարունակական նորացման կամ վերականգնման գործընթացը դրանց վնասումից հետո. Այն հյուսվածքներում, որոնք կայուն բջիջների պոպուլյացիաներ են և զուրկ են կամբիալ տարրերից (նյարդային հյուսվածք, սրտի մկանային հյուսվածք), վերականգնման այս տեսակը դրանց կառուցվածքն ու գործառույթը թարմացնելու և վերականգնելու միակ հնարավոր միջոցն է:
հյուսվածքների հիպերտրոֆիա- դրա ծավալի, զանգվածի և ֆունկցիոնալ գործունեության աճը սովորաբար հետևանք է ա) բջջային հիպերտրոֆիա(դրանց քանակով անփոփոխ) ուժեղացված ներբջջային ռեգեներացիայի շնորհիվ. բ) հիպերպլազիա -նրա բջիջների քանակի ավելացում՝ ակտիվացնելով բջիջների բաժանումը ( տարածում) և (կամ) նոր ձևավորված բջիջների տարբերակման արագացման արդյունքում. գ) երկու գործընթացների համակցություններ: հյուսվածքների ատրոֆիա- նրա ծավալի, զանգվածի և ֆունկցիոնալ ակտիվության նվազում՝ պայմանավորված ա) առանձին բջիջների ատրոֆիայի պատճառով՝ կատաբոլիզմի պրոցեսների գերակշռությամբ, բ) նրա որոշ բջիջների մահով, գ) բջիջների բաժանման արագության կտրուկ նվազմամբ և տարբերակում.
5. Միջհյուսվածքային և միջբջջային հարաբերություններ. Հյուսվածքը պահպանում է իր կառուցվածքային և ֆունկցիոնալ կազմակերպման կայունությունը (հոմեոստազ) որպես մեկ ամբողջություն միայն միմյանց վրա հիստոլոգիական տարրերի մշտական ազդեցության ներքո (ինտերստիցիալ փոխազդեցություններ), ինչպես նաև մեկ հյուսվածքի վրա (միջհյուսվածքային փոխազդեցություններ): Այդ ազդեցությունները կարելի է դիտարկել որպես տարրերի փոխադարձ ճանաչման, նրանց միջև շփումների ձևավորման և տեղեկատվության փոխանակման գործընթացներ։ Այս դեպքում ձևավորվում են կառուցվածքային-տարածական միավորումների բազմազանություն։ Հյուսվածքի բջիջները կարող են լինել հեռավորության վրա և փոխազդել միմյանց հետ միջբջջային նյութի միջոցով (միացնող հյուսվածքներ), շփվել գործընթացների հետ՝ երբեմն հասնելով զգալի երկարության (նյարդային հյուսվածք) կամ ձևավորել սերտորեն շփվող բջիջների շերտեր (էպիթելի): Հյուսվածքների ամբողջությունը շարակցական հյուսվածքի միջոցով միավորված մեկ կառուցվածքային ամբողջության մեջ, որի համակարգված գործունեությունը ապահովվում է նյարդային և հումորային գործոններով, ձևավորում է ամբողջ օրգանիզմի օրգաններ և օրգան համակարգեր։
Հյուսվածքի ձևավորման համար անհրաժեշտ է, որ բջիջները միավորվեն և փոխկապակցվեն բջջային անսամբլներում։ Բջիջների՝ միմյանց կամ միջբջջային նյութի բաղադրիչներին ընտրողաբար միանալու ունակությունն իրականացվում է ճանաչման և կպչման գործընթացների միջոցով, որոնք անհրաժեշտ պայման են հյուսվածքային կառուցվածքի պահպանման համար։ Ճանաչման և սոսնձման ռեակցիաները տեղի են ունենում հատուկ թաղանթային գլիկոպրոտեինների մակրոմոլեկուլների փոխազդեցության արդյունքում, որը կոչվում է. կպչուն մոլեկուլներ. Կցումը տեղի է ունենում հատուկ ենթաբջջային կառուցվածքների օգնությամբ՝ ա ) կետային կպչուն կոնտակտներ(բջիջների կցումը միջբջջային նյութին), բ) միջբջջային կապեր(բջիջների կցումը միմյանց):
Միջբջջային կապեր- բջիջների մասնագիտացված կառուցվածքներ, որոնց օգնությամբ դրանք մեխանիկորեն ամրացվում են միմյանց հետ, ինչպես նաև ստեղծում են միջբջջային հաղորդակցության խոչընդոտներ և թափանցելիության ուղիներ: Տարբերակել: 1) կպչուն բջիջների միացումներ, կատարելով միջբջջային կպչունություն (միջանկյալ կոնտակտ, դեզմոսոմ, կիսադեսմազոմ), 2) կապեր հաստատել, որի գործառույթը պատնեշի ձևավորումն է, որը թակարդում է նույնիսկ փոքր մոլեկուլները (սերտ շփում), 3) հաղորդիչ (հաղորդակցական) շփումներ, որի գործառույթն է ազդակներ փոխանցել բջջից բջիջ (բացի միացում, սինապս)։
6. Հյուսվածքների կենսագործունեության կարգավորում. Հյուսվածքների կարգավորումը հիմնված է երեք համակարգերի վրա՝ նյարդային, էնդոկրին և իմունային։ Հումորային գործոնները, որոնք ապահովում են միջբջջային փոխազդեցությունը հյուսվածքներում և դրանց նյութափոխանակությունը, ներառում են մի շարք բջջային մետաբոլիտներ, հորմոններ, միջնորդներ, ինչպես նաև ցիտոկիններ և խալոններ:
Ցիտոկիններ ներ և միջքաղաքային կարգավորող նյութերի առավել բազմակողմանի դասակարգն են: Դրանք գլիկոպրոտեիններ են, որոնք շատ ցածր կոնցենտրացիաների դեպքում ազդում են բջիջների աճի, բազմացման և տարբերակման ռեակցիաների վրա: Ցիտոկինների գործողությունը պայմանավորված է թիրախային բջիջների պլազմոլեմայի վրա նրանց համար ընկալիչների առկայությամբ։ Այս նյութերը կրում են արյունով և ունեն հեռավոր (էնդոկրին) ազդեցություն, ինչպես նաև տարածվում են միջբջջային նյութի միջով և գործում են տեղային (ավտո- կամ պարակրին): Ամենակարևոր ցիտոկիններն են ինտերլեյկիններ(IL), աճի գործոններ, գաղութը խթանող գործոններ(KSF), ուռուցքային նեկրոզ գործոն(TNF), ինտերֆերոն. Տարբեր հյուսվածքների բջիջները ունեն մեծ թվով ընկալիչներ տարբեր ցիտոկինների համար (10-ից մինչև 10000 մեկ բջջի համար), որոնց ազդեցությունները հաճախ համընկնում են, ինչը ապահովում է ներբջջային կարգավորման այս համակարգի գործունեության բարձր հուսալիություն:
Keylons- բջիջների բազմացման հորմոնանման կարգավորիչներ. արգելակում են միտոզը և խթանում բջիջների տարբերակումը: Քեյլոնները գործում են հետադարձ կապի սկզբունքով. հասուն բջիջների քանակի նվազմամբ (օրինակ՝ էպիդերմիսի կորուստ վնասվածքի պատճառով), քեյոնների թիվը նվազում է, և վատ տարբերակված կամբիալ բջիջների բաժանումը մեծանում է, ինչը հանգեցնում է հյուսվածքի: վերածնում.
Ուղարկել ձեր լավ աշխատանքը գիտելիքների բազայում պարզ է: Օգտագործեք ստորև բերված ձևը
Ուսանողները, ասպիրանտները, երիտասարդ գիտնականները, ովքեր օգտագործում են գիտելիքների բազան իրենց ուսումնառության և աշխատանքի մեջ, շատ շնորհակալ կլինեն ձեզ:
Տեղակայված է http://www.allbest.ru/ կայքում
Բելառուսի Հանրապետության գյուղատնտեսության և պարենի նախարարություն
Կրթական հաստատություն «Պատվո նշան Վիտեբսկի շքանշան»
Անասնաբուժության պետական ակադեմիա»
Պաթոլոգիական անատոմիայի և հյուսվածքաբանության ամբիոն
ԴԻՊԼՈՄԱՇԽԱՏԱՆՔ
«Ցիտոլոգիայի, հյուսվածաբանության և սաղմնաբանության հիմնախնդիրների ուսումնասիրություն» թեմայով.
Վիտեբսկ 2011 թ
1. Հյուսվածքաբանությունը որպես գիտություն, նրա հարաբերությունները այլ առարկաների հետ, դերը անասնաբույժի ձևավորման և գործնական աշխատանքի մեջ.
2. «Բջջ» հասկացության սահմանում. Նրա կառուցվածքային կազմակերպումը
3. Ցիտոպլազմայի կազմը և նպատակը
4. Բջջային օրգանելներ (սահմանում, դասակարգում, միտոքոնդրիաների կառուցվածքի և գործառույթների բնութագրում, շերտավոր համալիր, լիզոսոմներ, էնդոպլազմային ցանց)
5. Միջուկի կառուցվածքը և գործառույթները
6. Բջջային բաժանման տեսակները
8. Սպերմատոզոիդների կառուցվածքը և նրանց կենսաբանական հատկությունները
9. Սպերմատոգենեզ
10. Ձվերի կառուցվածքը և դասակարգումը
11. Սաղմի զարգացման փուլերը
12. Կաթնասունների սաղմնային զարգացման առանձնահատկությունները (տրոֆոբլաստների և պտղի թաղանթների ձևավորում)
13. Պլասենտա (կառուցվածք, գործառույթներ, դասակարգումներ)
14. Մորֆոլոգիական դասակարգում և -ի համառոտ նկարագրությունըէպիթելի հիմնական տեսակները
15. ընդհանուր բնութագրերըարյունը որպես մարմնի ներքին միջավայրի հյուսվածք
16. Գրանուլոցիտների կառուցվածքը և ֆունկցիոնալ նշանակությունը
17. Ագրանուլոցիտների կառուցվածքը և ֆունկցիոնալ նշանակությունը
18. Չամրացված շարակցական հյուսվածքի մորֆոֆունկցիոնալ բնութագրերը
19. Նյարդային հյուսվածքի ընդհանուր բնութագրերը (նեյրոցիտների և նեյրոգլիայի կազմը, դասակարգումը)
20. Տիմուսի կառուցվածքը և գործառույթները
21. Լիմֆյան հանգույցների կառուցվածքը և գործառույթները
22. Կառուցվածքը և գործառույթները
23. Միապալատ ստամոքսի կառուցվածքն ու գործառույթները. Նրա մռայլ ապարատի առանձնահատկությունները
24. Բարակ աղիքի կառուցվածքը և գործառույթները
25. Լյարդի կառուցվածքը և գործառույթները
26. Թոքերի կառուցվածքը և գործառույթները
27. Երիկամների կառուցվածքը և գործառույթները
28. Ամորձիների կառուցվածքը և գործառույթները
29. Արգանդի կառուցվածքն ու գործառույթները
30. Էնդոկրին համակարգի կազմը և նպատակը
31. Ուղեղի կեղեւի բջջային կառուցվածքը
1. Գհյուսվածաբանությունը որպես գիտություն, նրա հարաբերությունները այլ առարկաների հետ, դերը անասնաբուժական բժշկի ձևավորման և գործնական աշխատանքի մեջ.
Հիստոլոգիան (histos - հյուսվածք, logos - ուսուցում, գիտություն) գիտություն է կենդանիների և մարդկանց բջիջների, հյուսվածքների և օրգանների մանրադիտակային կառուցվածքի, զարգացման և կենսագործունեության մասին։ Մարմինը միասնական ամբողջական համակարգ է, որը կառուցված է բազմաթիվ մասերից: Այս մասերը սերտորեն փոխկապակցված են, և օրգանիզմն ինքը մշտապես փոխազդում է արտաքին միջավայրի հետ։ Էվոլյուցիայի գործընթացում կենդանական օրգանիզմը ձեռք է բերել իր կազմակերպման բազմաստիճան բնույթ.
Մոլեկուլային.
Ենթաբջջային.
Բջջային.
Հյուսվածք.
Օրգան.
Համակարգ.
Օրգանական.
Սա թույլ է տալիս կենդանիների կառուցվածքն ուսումնասիրելիս նրանց օրգանիզմները բաժանել առանձին մասերի, կիրառել հետազոտության տարբեր մեթոդներ և որպես գիտելիքի առանձին ճյուղ առանձնացնել հյուսվածաբանության հետևյալ բաժինները.
1. Ցիտոլոգիա - ուսումնասիրում է մարմնի բջիջների կառուցվածքն ու գործառույթները;
2. Սաղմնաբանություն - ուսումնասիրում է մարմնի սաղմնային զարգացման օրինաչափությունները.
ա) ընդհանուր սաղմնաբանություն՝ սաղմի զարգացման ամենավաղ փուլերի գիտություն, ներառյալ օրգանների առաջացման շրջանը, որոնք բնութագրում են անհատների պատկանելությունը կենդանական թագավորության որոշակի տեսակի և դասի.
բ) մասնավոր սաղմնաբանություն՝ սաղմի բոլոր օրգանների և հյուսվածքների զարգացման վերաբերյալ գիտելիքների համակարգ.
3. Ընդհանուր հիստոլոգիա - մարմնի հյուսվածքների կառուցվածքի և ֆունկցիոնալ հատկությունների ուսումնասիրություն;
4. Մասնավոր հյուսվածաբանություն - կարգապահության առավել ընդարձակ և կարևոր բաժին, ներառյալ մարմնի որոշակի համակարգեր ձևավորող օրգանների կառուցվածքային առանձնահատկությունների և ֆունկցիոնալ գործառույթների վերաբերյալ գիտելիքների ամբողջականությունը:
Հյուսվածքաբանությունը պատկանում է մորֆոլոգիական գիտություններին և կենսաբանական հիմնարար գիտություններից է։ Այն սերտորեն կապված է այլ ընդհանուր կենսաբանության հետ (կենսաքիմիա, անատոմիա, գենետիկա, ֆիզիոլոգիա, իմունոմորֆոլոգիա, մոլեկուլային կենսաբանություն), անասնաբուծական համալիրի դիսցիպլինները, ինչպես նաև անասնաբուժական պրոֆիլը (ախտաբանական անատոմիա, անասնաբուժասանիտարական հետազոտություն, մանկաբարձություն, թերապիա և այլն): Նրանք միասին կազմում են անասնաբուժության ուսումնասիրության տեսական հիմքը: Հյուսվածքաբանությունը նաև մեծ գործնական նշանակություն ունի՝ բժշկական պրակտիկայում լայնորեն կիրառվում են բազմաթիվ հյուսվածաբանական հետազոտական մեթոդներ։
Հյուսվածքաբանության առաջադրանքներն ու նշանակությունը.
1. Այլ գիտությունների հետ կազմում է բժշկական մտածողություն։
2. Հյուսվածքաբանությունը կենսաբանական հիմք է ստեղծում անասնաբուժության և անասնաբուծության զարգացման համար:
3. Կենդանիների հիվանդությունների ախտորոշման ժամանակ լայնորեն կիրառվում են հյուսվածաբանական մեթոդները։
4. Հյուսվածքաբանությունը ապահովում է կերային հավելումների և պրոֆիլակտիկ միջոցների օգտագործման որակի և արդյունավետության հսկողություն:
5. Հյուսվածքաբանական հետազոտության մեթոդների օգնությամբ վերահսկվում է անասնաբուժական պատրաստուկների թերապեւտիկ արդյունավետությունը։
6. Տրամադրում է կենդանիների հետ բուծման աշխատանքների որակի եւ հոտի վերարտադրության գնահատականը.
7. Կենդանիների օրգանիզմում ցանկացած նպատակային միջամտություն կարելի է վերահսկել հյուսվածքաբանական մեթոդներով։
2. «Բջջ» տերմինի սահմանումը. Նրա կառուցվածքային կազմակերպումը
Բջիջը հիմնական կառուցվածքային և ֆունկցիոնալ միավորն է, որը ընկած է կենդանական և բուսական օրգանիզմների կառուցվածքի, զարգացման և կյանքի հիմքում: Այն բաղկացած է 2 անքակտելիորեն կապված մասերից՝ ցիտոպլազմից և միջուկից։ Ցիտոպլազմը ներառում է 4 բաղադրիչ.
բջջային պատ (պլազմոլեմա):
Հիալոպլազմ
Օրգանելներ (օրգանելներ)
Բջջային ընդգրկումներ
Միջուկը նույնպես բաղկացած է 4 մասից.
Միջուկային թաղանթ կամ կարիոլեմա
Միջուկային հյութ կամ կարիոպլազմ
Քրոմատին
Պլազմալեման բջջի արտաքին թաղանթն է։ Այն կառուցված է կենսաբանական թաղանթից, վերթաղանթային համալիրից և ենթաթաղանթային ապարատից։ Պահում է բջջային պարունակությունը, պաշտպանում է բջիջը և ապահովում դրա փոխազդեցությունը պերիբջջային միջավայրի, այլ բջիջների և հյուսվածքային տարրերի հետ:
Հիալոպլազմը ցիտոպլազմայի կոլոիդային միջավայր է: Ծառայում է օրգանելների, ներդիրների տեղադրման, դրանց փոխազդեցության իրականացման համար։
Օրգանելները ցիտոպլազմայի մշտական կառուցվածքներն են, որոնք որոշակի գործառույթներ են կատարում դրանում։
Ներառումներ - նյութեր, որոնք մտնում են բջիջ սննդային նպատակներով կամ գոյանում են դրանում կենսական գործընթացների արդյունքում։
Միջուկային թաղանթը բաղկացած է երկու կենսաբանական թաղանթներից, սահմանազատում է միջուկի պարունակությունը ցիտոպլազմայից և միևնույն ժամանակ ապահովում դրանց սերտ փոխազդեցությունը։
Միջուկային հյութը միջուկի կոլոիդային միջավայր է։
Քրոմատինը քրոմոսոմների գոյության ձև է։ Բաղկացած է ԴՆԹ-ից, հիստոնային և ոչ հիստոնային սպիտակուցներից՝ ՌՆԹ-ից։
Նուկլեոլուսը միջուկային կազմակերպիչների, ռիբոսոմային ՌՆԹ-ի, սպիտակուցների և ռիբոսոմների ստորաբաժանումների ԴՆԹ-ի համալիր է, որոնք ձևավորվում են այստեղ։
3. Ցիտոպլազմայի կազմը և նպատակը
Ցիտոպլազմը բջջի երկու հիմնական մասերից մեկն է, որն ապահովում է նրա կյանքի հիմնական գործընթացները։
Ցիտոպլազմը ներառում է 4 բաղադրիչ.
Բջջային թաղանթ (պլազմոլեմմա):
Հիալոպլազմ.
Օրգանելներ (օրգանելներ):
Բջջային ընդգրկումներ.
Հիալոպլազմը ցիտոպլազմայի կոլոիդային մատրիցն է, որում տեղի են ունենում բջջի կենսագործունեության հիմնական պրոցեսները, տեղակայվում և գործում են օրգանելները և ներդիրները։
Բջջային թաղանթը (պլազմոլեմմա) կառուցված է կենսաբանական թաղանթից, վերթաղանթային համալիրից և ենթաթաղանթային ապարատից։ Այն պահպանում է բջջային պարունակությունը, պահպանում է բջիջների ձևը, իրականացնում է նրանց շարժիչային ռեակցիաները, կատարում է արգելքների և ընկալիչների գործառույթներ, ապահովում է նյութերի ընդունման և արտազատման գործընթացները, ինչպես նաև փոխազդեցությունը պերիբջջային միջավայրի, այլ բջիջների և հյուսվածքային տարրերի հետ:
Կենսաբանական թաղանթը՝ որպես պլազմոլեմմայի հիմք, կառուցված է երկմոլեկուլային լիպիդային շերտից, որի մեջ խճանկարային կերպով ներառված են սպիտակուցի մոլեկուլները։ Լիպիդային մոլեկուլների հիդրոֆոբ բևեռները շրջված են դեպի ներս՝ ձևավորելով մի տեսակ հիդրավլիկ կողպեք, իսկ դրանց հիդրոֆիլ գլուխները ապահովում են ակտիվ փոխազդեցություն արտաքին և ներբջջային միջավայրի հետ։
Սպիտակուցները գտնվում են մակերեսորեն (ծայրամասային), մտնում են հիդրոֆոբ շերտը (կիսաինտեգրալ) կամ թաղանթում ներթափանցում (ինտեգրալ): Ֆունկցիոնալ առումով նրանք կազմում են կառուցվածքային, ֆերմենտային, ընկալիչ և փոխադրող սպիտակուցներ։
Գլիկոզամինոգլիկանների կողմից ձևավորվում է վերմեմբրանային համալիր՝ գլիկոկալիքս, թաղանթներ։ Կատարում է պաշտպանիչ և կարգավորող գործառույթներ:
Ենթաթաղանթային ապարատը ձևավորվում է միկրոխողովակներով և միկրոթելերով: Գործում է որպես մկանային-կմախքային համակարգ:
Օրգանելները ցիտոպլազմայի մշտական կառուցվածքներն են, որոնք որոշակի գործառույթներ են կատարում դրանում։ Կան ընդհանուր նշանակության օրգանելներ (Գոլջիի ապարատ, միտոքոնդրիա, բջջային կենտրոն, ռիբոսոմներ, լիզոսոմներ, պերօքսիսոմներ, ցիտոպլազմային ցանց, միկրոխողովակներ և միկրոթելեր) և հատուկ (միոֆիբրիլներ՝ մկանային բջիջներում; նեյրոֆիբրիլներ, սինապտիկ վեզիկուլներ և վագրային նյութ՝ նեյրոֆիբրիլներ, միկրոֆիբրիլներ, նեյրոֆիբրիլներ. , թարթիչ և դրոշակ - էպիթելային բջիջներում):
Ներառումներ - նյութեր, որոնք մտնում են բջիջ սննդային նպատակներով կամ գոյանում են դրանում կենսական գործընթացների արդյունքում։ Տարբերում են տրոֆիկ, սեկրետորային, պիգմենտային և արտազատվող ներդիրներ։
4. Բջջային օրգանելներ (սահմանում, դասակարգում, միտոքոնդրիաների կառուցվածքի և գործառույթների բնութագրում, շերտավոր համալիր, լիզոսոմներ, էնդոպլազմիկ ցանց)
Օրգանելները (օրգանելներ) ցիտոպլազմայի մշտական կառուցվածքներն են, որոնք որոշակի գործառույթներ են կատարում դրանում։
Օրգանելների դասակարգումը հաշվի է առնում նրանց կառուցվածքի և ֆիզիոլոգիական գործառույթների առանձնահատկությունները:
Կատարված գործառույթների բնույթից ելնելով բոլոր օրգանելները բաժանվում են երկու մեծ խմբերի.
1. Ընդհանուր նշանակության օրգանները՝ արտահայտված մարմնի բոլոր բջիջներում, ապահովում են դրանց կառուցվածքի և կենսագործունեության ապահովման ամենատարածված գործառույթները (միտոքոնդրիաներ, ցենտրոսոմներ, ռիբոսոմներ, լիզոսոմներ, պերօքսիսոմներ, միկրոխողովակներ, ցիտոպլազմային ցանց, Գոլջիի համալիր)
2. Հատուկ - հայտնաբերված է միայն հատուկ գործառույթներ կատարող բջիջներում (միոֆիբրիլներ, տոնոֆիբրիլներ, նեյրոֆիբրիլներ, սինապտիկ վեզիկուլներ, տիգրոիդ նյութ, միկրովիլիներ, թարթիչներ, դրոշակներ):
Ըստ կառուցվածքային առանձնահատկությունի՝ առանձնացնում ենք թաղանթային և ոչ թաղանթային կառուցվածքի օրգանելներ։
Մեմբրանային օրգանելները հիմնականում ունեն մեկ կամ երկու կենսաբանական թաղանթ (միտոքոնդրիա, շերտավոր բարդույթ, լիզոսոմներ, պերօքսիսոմներ, էնդոպլազմիկ ցանց):
Ոչ թաղանթային օրգանելները ձևավորվում են միկրոխողովակներով՝ մոլեկուլների համալիրից և դրանց կապոցներից (ցենտրոսոմներ, միկրոխողովակներ, միկրոթելեր և ռիբոսոմներ) գլոբուլներով։
Ըստ չափի՝ մենք առանձնացնում ենք լույսի մանրադիտակով տեսանելի օրգանելների խումբ (Գոլջիի ապարատ, միտոքոնդրիա, բջջային կենտրոն), և ուլտրամիկրոսկոպիկ օրգանելներ՝ տեսանելի միայն էլեկտրոնային մանրադիտակի տակ (լիզոսոմներ, պերօքսիսոմներ, ռիբոսոմներ, էնդոպլազմիկ ցանց, միկրոխողովակներ և միկրոթելեր):
Գոլջիի համալիրը (շերտավոր բարդույթ) տեսանելի է լուսային մանրադիտակի տակ՝ կարճ և երկար թելերի տեսքով (մինչև 15 մկմ երկարությամբ): Էլեկտրոնային մանրադիտակով յուրաքանչյուր նման թել (դիկտյոսոմ) իրենից ներկայացնում է միմյանց վրա շերտավորված հարթ ցիստեռնների, խողովակների և վեզիկուլների համալիր: Շերտավոր համալիրն ապահովում է գաղտնիքների կուտակումն ու արտազատումը, սինթեզում է որոշ լիպիդներ և ածխաջրեր, ձևավորում առաջնային լիզոսոմներ։
Լույսի մանրադիտակի տակ գտնվող միտոքոնդրիաները հայտնաբերվում են բջիջների ցիտոպլազմայում մանր հատիկների և կարճ թելերի տեսքով (մինչև 10 մկմ երկարությամբ), որոնց անուններից էլ ձևավորվում է օրգանոիդի հենց անվանումը։ Էլեկտրոնային մանրադիտակով նրանցից յուրաքանչյուրը հայտնվում է կլոր կամ երկարավուն մարմինների տեսքով՝ բաղկացած երկու թաղանթից և մատրիցից։ Ներքին թաղանթն ունի սրածայրման ելուստներ՝ cristae։ Մատրիցը պարունակում է միտոքոնդրիալ ԴՆԹ և ռիբոսոմներ, որոնք սինթեզում են որոշ կառուցվածքային սպիտակուցներ։ Միտոքոնդրիալ թաղանթների վրա տեղայնացված ֆերմենտներն ապահովում են օրգանական նյութերի օքսիդացման գործընթացները (բջջային շնչառություն) և ATP-ի պահպանումը (էներգիայի ֆունկցիա):
Լիզոսոմները ներկայացված են փոքրիկ պղպջակների նման գոյացություններով, որոնց պատը կազմված է կենսաբանական թաղանթով, որի ներսում պարփակված է հիդրոլիտիկ ֆերմենտների լայն տեսականի (մոտ 70)։
Նրանք խաղում են բջիջների մարսողական համակարգի դերը, չեզոքացնում են վնասակար նյութերը և օտար մասնիկները և օգտագործում իրենց հնացած և վնասված կառուցվածքները:
Տարբերում են առաջնային լիզոսոմներ, երկրորդային (ֆագոլիզոսոմներ, աուտոֆագոլիզոսոմներ) և երրորդական տելոլիզոսոմներ (մնացորդային մարմիններ)։
Էնդոպլազմիկ ցանցը փոքրիկ տանկերի և խողովակների համակարգ է, որոնք անաստոմոզ են միմյանց հետ և ներթափանցում ցիտոպլազմա: Նրանց պատերը ձևավորվում են միայնակ թաղանթներով, որոնց վրա պատվիրված են լիպիդների և ածխաջրերի սինթեզի ֆերմենտներ՝ հարթ էնդոպլազմիկ ցանց (ագրանուլյար) կամ ամրագրված են ռիբոսոմներ՝ կոպիտ (հատիկավոր) ցանց։ Վերջինս նախատեսված է օրգանիզմի ընդհանուր կարիքների համար սպիտակուցի մոլեկուլների արագացված սինթեզի համար (արտահանման համար)։ EPS-ի երկու տեսակներն էլ ապահովում են տարբեր նյութերի շրջանառություն և տեղափոխում:
անասնաբուժական բժշկության հյուսվածքաբանություն բջջային օրգանիզմ
5. Միջուկի կառուցվածքը և գործառույթները
Բջջի միջուկը նրա երկրորդ կարևոր բաղադրիչն է:
Բջիջների մեծ մասն ունի մեկ միջուկ, սակայն լյարդի որոշ բջիջներ և կարդիոմիոցիտներ ունեն 2 միջուկ: Ոսկրային հյուսվածքի մակրոֆագներում դրանք 3-ից մի քանի տասնյակ են, իսկ գծավոր մկանային մանրաթելում՝ 100-ից 3 հազար միջուկ։ Ընդհակառակը, կաթնասունների էրիթրոցիտները միջուկային չեն:
Միջուկի ձևը հաճախ կլորացված է, բայց էպիթելի պրիզմատիկ բջիջներում այն ձվաձև է, հարթ բջիջներում՝ հարթեցված, հասուն հատիկավոր լեյկոցիտներում՝ հատվածավորված, հարթ միոցիտներում երկարանում է մինչև ձողաձև։ Միջուկը, որպես կանոն, գտնվում է բջջի կենտրոնում։ Պլազմային բջիջներում այն ընկած է էքսցենտրիկ կերպով, իսկ պրիզմատիկ էպիթելային բջիջներում այն տեղափոխվում է բազալ բևեռ:
Միջուկի քիմիական կազմը.
սպիտակուցներ՝ 70%, նուկլեինաթթուներ՝ 25%, ածխաջրեր, լիպիդներ և անօրգանական նյութերկազմում են մոտավորապես 5%:
Կառուցվածքային առումով միջուկը կառուցված է.
1. միջուկային թաղանթ (կարիոլեմմա),
2. միջուկային հյութ (կարիոպլազմ),
3. միջուկ,
4. քրոմատին Միջուկային թաղանթ՝ կարիոլեմման բաղկացած է 2 տարրական կենսաբանական թաղանթից։ Նրանց միջև արտահայտված է պերինուկլեար տարածություն։ Որոշ հատվածներում երկու թաղանթները փոխկապակցված են և կազմում են կարիոլեմայի ծակոտիներ՝ մինչև 90 նմ տրամագծով։ Նրանք ունեն կառուցվածքներ, որոնք կազմում են, այսպես կոչված, երեք թիթեղների ծակոտիների համալիրը: Յուրաքանչյուր ափսեի եզրերի երկայնքով կա 8 հատ, իսկ կենտրոնում՝ մեկը։ Ամենաբարակ մանրաթելերը (թելերը) դրան են գնում ծայրամասային հատիկներից։ Արդյունքում ձևավորվում են յուրահատուկ դիֆրագմներ, որոնք կարգավորում են օրգանական մոլեկուլների և դրանց բարդույթների շարժումը պատյանով։
Karyolemma գործառույթները:
1. սահմանազատող,
2. կարգավորող.
Միջուկային հյութը (կարիոպլազմ) ածխաջրերի, սպիտակուցների, նուկլեոտիդների և հանքանյութերի կոլոիդային լուծույթ է։ Այն միկրոմիջավայր է նյութափոխանակության ռեակցիաներ ապահովելու և սուրհանդակի և ՌՆԹ-ի տեղափոխման համար միջուկային ծակոտիներ:
Քրոմատինը քրոմոսոմների գոյության ձև է։ Այն ներկայացված է ԴՆԹ, ՌՆԹ մոլեկուլների, փաթեթավորող սպիտակուցների և ֆերմենտների (հիստոններ և ոչ հիստոնային սպիտակուցներ) համալիրով։ Հիստոնները ուղղակիորեն կապված են քրոմոսոմին: Նրանք ապահովում են քրոմոսոմում ԴՆԹ-ի մոլեկուլի պարույրացումը։ Ոչ հիստոնային սպիտակուցները ֆերմենտներ են. ԴՆԹ - նուկլեազներ, որոնք ոչնչացնում են կոմպլեմենտար կապերը՝ առաջացնելով դրա դեսպիրալացում;
ԴՆԹ և ՌՆԹ - պոլիմերազներ, որոնք ապահովում են ՌՆԹ-ի մոլեկուլների կառուցումը ասեղնագործված ԴՆԹ-ի վրա, ինչպես նաև քրոմոսոմների ինքնակրկնօրինակումը մինչև բաժանումը։
Քրոմատինը միջուկում առկա է երկու ձևով.
1. ցրված էխրոմատին, որն արտահայտվում է նուրբ հատիկների և թելերի տեսքով։ Այս դեպքում ԴՆԹ-ի մոլեկուլների հատվածները գտնվում են չոլորված վիճակում: Դրանց վրա ՌՆԹ-ի մոլեկուլները հեշտությամբ սինթեզվում են, սպիտակուցի կառուցվածքի մասին տեղեկություններ կարդալով, կառուցվում են տրանսպորտային ՌՆԹ-ներ։ Ստացված և - ՌՆԹ-ն տեղափոխվում է ցիտոպլազմա և ներմուծվում ռիբոսոմների մեջ, որտեղ իրականացվում են սպիտակուցների սինթեզի գործընթացները։ Euchromatin-ը քրոմատինի ֆունկցիոնալ ակտիվ ձևն է: Նրա գերակշռությունը վկայում է բջիջների կենսական գործընթացների բարձր մակարդակի մասին։
2. Խտացրած հետերոքրոմատին. Լույսի մանրադիտակի տակ այն նման է խոշոր հատիկների և կուտակումների: Միևնույն ժամանակ, հիստոնային սպիտակուցները սերտորեն ոլորում և փաթեթավորում են ԴՆԹ-ի մոլեկուլները, որոնց վրա, հետևաբար, անհնար է կառուցել և - ՌՆԹ, այդ իսկ պատճառով հետերոքրոմատինը քրոմոսոմային հավաքածուի ֆունկցիոնալ ոչ ակտիվ, չպահանջված մասն է:
Միջուկ. Այն ունի կլորացված ձև՝ մինչև 5 մկմ տրամագծով։ Բջիջներում կարող են արտահայտվել 1-ից 3 միջուկներ՝ կախված դրա ֆունկցիոնալ վիճակից։ Ներկայացնում է մի քանի քրոմոսոմների տերմինալային հատվածների մի շարք, որոնք կոչվում են միջուկային կազմակերպիչներ։ Միջուկային կազմակերպիչների ԴՆԹ-ի վրա առաջանում են ռիբոսոմային ՌՆԹ-ներ, որոնք համապատասխան սպիտակուցների հետ միանալիս կազմում են ռիբոսոմային ենթամիավորներ։
Միջուկի գործառույթները.
1. Մայր բջջից ստացված ժառանգական տեղեկատվության պահպանումն անփոփոխ.
2. Կենսական գործընթացների համակարգումը և ժառանգական տեղեկատվության ներդրումը կառուցվածքային և կարգավորող սպիտակուցների սինթեզի միջոցով:
3. Բաժանման ժամանակ ժառանգական տեղեկատվության փոխանցումը դուստր բջիջներին:
6. Բջիջների բաժանման տեսակները
Բաժանումը բջիջների ինքնավերարտադրման միջոց է։ Այն ապահովում է.
ա) որոշակի տեսակի բջիջների գոյության շարունակականությունը.
բ) հյուսվածքների հոմեոստազ;
գ) հյուսվածքների և օրգանների ֆիզիոլոգիական և վերականգնողական վերականգնում.
դ) անհատների բազմացում և կենդանիների տեսակների պահպանում.
Բջիջների բաժանման 3 եղանակ կա.
1. ամիտոզ - բջիջների բաժանում առանց քրոմոսոմային ապարատի տեսանելի փոփոխությունների: Այն առաջանում է միջուկի և ցիտոպլազմայի պարզ սեղմումով։ Քրոմոսոմները չեն հայտնաբերվում, բաժանման spindle չի ձևավորվում: Բնորոշ է սաղմնային և վնասված որոշ հյուսվածքների համար։
2. միտոզ - վերարտադրության փուլում սոմատիկ և սեռական բջիջների բաժանման մեթոդ: Այս դեպքում մեկ մայր բջջից ձևավորվում են քրոմոսոմների ամբողջական կամ դիպլոիդ բջիջներով երկու դուստր բջիջ։
3. Մեյոզը հասունացման փուլում սեռական բջիջների բաժանման մեթոդ է, որի դեպքում մեկ մայր բջջից ձևավորվում են 4 դուստր բջիջներ՝ կիսով չափ, հապլոիդ, քրոմոսոմներով։
7. Միտոզ
Միտոզին նախորդում է ինտերֆազը, որի ընթացքում բջիջը պատրաստվում է ապագա բաժանմանը։ Այս թրեյնինգը ներառում է
բջիջների աճ;
Էներգիայի պահպանում ATP-ի և սննդանյութերի տեսքով;
ԴՆԹ-ի մոլեկուլների և քրոմոսոմային հավաքածուի ինքնակրկնապատկում: Կրկնապատկման արդյունքում յուրաքանչյուր քրոմոսոմ բաղկացած է 2 քույր քրոմատիդներից;
Բջջային կենտրոնի ցենտրիոլների կրկնապատկում;
Հատուկ սպիտակուցների սինթեզ, ինչպիսին է տուբուլինը, տրոհվող լիսեռի թելեր ստեղծելու համար:
Միտոզը ինքնին բաղկացած է 4 փուլից.
պրոֆազ,
մետաֆազ,
անաֆազ,
Տելոֆազ.
Պրոֆազում քրոմոսոմները գալարվում են, խտանում և կարճանում։ Նրանք այժմ տեսանելի են լուսային մանրադիտակի տակ: Բջջային կենտրոնի ցենտրիոլները սկսում են շեղվել դեպի բևեռները: Նրանց միջև կառուցված է բաժանման լիսեռ: Պրոֆազի վերջում միջուկը անհետանում է և տեղի է ունենում միջուկային ծրարի մասնատում։
Մետաֆազում ավարտված է բաժանման լիսեռի կառուցումը: Կարճ լիսեռ թելերը կցվում են քրոմոսոմների ցենտրոմերներին։ Բոլոր քրոմոսոմները գտնվում են բջջի հասարակածում: Նրանցից յուրաքանչյուրը պահվում է հասարակածային ափսեում 2 քրոմատինային թելերի օգնությամբ, որոնք գնում են դեպի բջջի բևեռները, և նրա կենտրոնական գոտին լցված է երկար ախրոմատինային մանրաթելերով։
Անաֆազում, քրոմատինային թելերի կծկման պատճառով, ցենտրոմերների շրջանում քրոմատիդների բաժանման spindles բաժանվում են միմյանցից, որից հետո նրանցից յուրաքանչյուրը սահում է կենտրոնական թելերի երկայնքով դեպի բջջի վերին կամ ստորին բևեռը: Այս պահից սկսած քրոմատիդը կոչվում է քրոմոսոմ: Այսպիսով, բջջի բևեռներում կա հավասար թվով միանման քրոմոսոմներ, այսինքն. դրանցից մեկ ամբողջական, դիպլոիդ հավաքածու։
Տելոֆազում քրոմոսոմների յուրաքանչյուր խմբի շուրջ ձևավորվում է նոր միջուկային ծրար։ Խտացրած քրոմատինը սկսում է թուլանալ։ Հայտնվում են նուկլեոլներ: Բջջի կենտրոնական մասում պլազմոլեմման դուրս է ցցվում դեպի ներս, դրան միացված են էնդոպլազմիկ ցանցի խողովակները, ինչը հանգեցնում է ցիտոտոմիայի և մայր բջջի բաժանմանը երկու դուստր բջիջների։
Մեյոզ (կրճատման բաժանում):
Դրան նախորդում է նաև ինտերֆազը, որում առանձնանում են նույն պրոցեսները, ինչ մինչ միտոզը։ Մեյոզն ինքնին ներառում է երկու բաժանում՝ կրճատում, որում ձևավորվում են կրկնակի քրոմոսոմներով հապլոիդ բջիջներ, և հավասարազոր, որը միտոզով հանգեցնում է միայնակ քրոմոսոմներով բջիջների ձևավորմանը։
Առաջատար երեւույթը, որն ապահովում է քրոմոսոմային հավաքածուի նվազումը, յուրաքանչյուր զույգում հայրական և մայրական քրոմոսոմների խոնարհումն է, որը տեղի է ունենում առաջին բաժանման պրոֆազում։ Երբ երկու քրոմատիդներից բաղկացած հոմոլոգ քրոմոսոմները մոտենում են միմյանց, ձևավորվում են տետրադներ, որոնք արդեն ներառում են 4 քրոմատիդներ։
Մեյոզի մետաֆազում տետրադները պահպանվում են և գտնվում են բջջի հասարակածում։ Անաֆազում, հետևաբար, ամբողջ կրկնապատկված քրոմոսոմները հեռանում են դեպի բևեռներ: Արդյունքում երկու դուստր բջիջ են ձևավորվում կրկնապատկված քրոմոսոմների կես հավաքածուով: Նման բջիջները շատ կարճ ինտերֆազից հետո նորից բաժանվում են նորմալ միտոզով, ինչը հանգեցնում է միայնակ քրոմոսոմներով հապլոիդ բջիջների առաջացմանը։
Հոմոլոգ քրոմոսոմների խոնարհման ֆենոմենը միաժամանակ լուծում է մեկ այլ կարևոր խնդիր՝ անհատական գենետիկ փոփոխականության նախադրյալների ստեղծում՝ կապված առաջին բաժանման մետաֆազում տետրադների բևեռային կողմնորոշման մեջ գեների փոխանակման և գեների փոխանակման գործընթացների հետ:
8. Սպերմատոզոիդների կառուցվածքը և դրանց կենսաբանական հատկությունները
Սպերմատոզոիդները (արական սեռական բջիջները) դրոշակավոր բջիջներ են։ Սպերմատոզոիդում օրգանելների հաջորդական դասավորությունը հնարավորություն է տալիս բջջում տարբերել գլուխը, պարանոցը, մարմինը և պոչը։
Գյուղատնտեսական կաթնասունների ներկայացուցիչների սպերմատոզոիդների գլուխը ասիմետրիկ է՝ դույլաձեւ, որն ապահովում է նրա ուղղագիծ, թարգմանական-պտտվող շարժումը։ Գլխի մեծ մասը զբաղեցնում է միջուկը, իսկ ամենաառաջնային մասը կազմում է գլխի գլխարկը՝ ակրոսոմով։ Ակրոսոմում (ձևափոխված Գոլջիի համալիր) կուտակվում են ֆերմենտներ (հիալուրոնիդազ, պրոթեզեր), որոնք թույլ են տալիս սերմնաբջիջներին բեղմնավորման ընթացքում ոչնչացնել ձվի երկրորդական թաղանթները։
Միջուկի հետևում, բջջի պարանոցում, մեկը մյուսի հետևից գտնվում են երկու ցենտրիոլներ՝ պրոքսիմալ և դիստալ։ Մոտակա ցենտրիոլն ազատորեն գտնվում է ցիտոպլազմայում և բեղմնավորման ընթացքում ներմուծվում է ձվի մեջ: Դիստալ ցենտրիոլից աճում է առանցքային թել. սա հատուկ բջջային օրգանել է, որն ապահովում է պոչի հարվածը միայն մեկ հարթությունում:
Սռնու թելի շուրջ սերմնահեղուկի մարմնում միտոքոնդրիումները գտնվում են մեկը մյուսի հետևից՝ կազմելով պարուրաձև թել՝ բջջի էներգետիկ կենտրոնը։
Պոչի շրջանում ցիտոպլազմը աստիճանաբար նվազում է, այնպես որ դրա վերջնական մասում առանցքային թելիկը հագցվում է միայն պլազմոլեմայի կողմից։
Սպերմատոզոիդների կենսաբանական հատկությունները.
1. Հայրական օրգանիզմի մասին ժառանգական տեղեկատվության կրելը.
2. Սպերմատոզոիդները բաժանման ընդունակ չեն, նրանց միջուկը պարունակում է քրոմոսոմների կես (հապլոիդ) հավաքածու։
3. Բջիջների չափը չի փոխկապակցվում կենդանիների քաշի հետ, և, հետևաբար, գյուղատնտեսական կաթնասունների ներկայացուցիչների մոտ այն տատանվում է նեղ սահմաններում (35-ից մինչև 63 միկրոն):
4. Շարժման արագությունը րոպեում 2-5մմ է։
5. Սպերմատոզոիդներին բնորոշ է ռեոտաքսիսի երեւույթը, այսինքն. շարժում կանանց սեռական տրակտում լորձի թույլ հոսանքի դեմ, ինչպես նաև քիմոտաքսիսի երևույթը` սերմնաբջիջների շարժումը դեպի քիմիկատներ (գինոգամոններ), որոնք արտադրվում են ձվի կողմից:
6. Էպիդիդիմիսում սպերմատոզոիդները ձեռք են բերում լրացուցիչ լիպոպրոտեինային ծածկույթ, որը թույլ է տալիս թաքցնել իրենց անտիգենները, քանի որ Կանանց մարմնի համար արական սեռական բջիջները գործում են որպես օտար բջիջներ:
7. Սպերմատոզոիդներն ունեն բացասական լիցք, որը նրանց հնարավորություն է տալիս վանելու միմյանց և դրանով իսկ կանխում բջիջների սոսնձումն ու մեխանիկական վնասումը (մեկ սերմնաժայթքում կա մինչև մի քանի միլիարդ բջիջ):
8. Ներքին բեղմնավորմամբ կենդանիների սպերմատոզոիդները չեն դիմանում շրջակա միջավայրի գործոնների ազդեցությանը, որի դեպքում նրանք մահանում են գրեթե անմիջապես։
9. Բարձր ջերմաստիճանը, ուլտրամանուշակագույն ճառագայթումը, թթվային միջավայրը, ծանր մետաղների աղերը վնասակար ազդեցություն են ունենում սպերմատոզոիդների վրա։
10. Անբարենպաստ ազդեցությունները դրսևորվում են ճառագայթման, ալկոհոլի, նիկոտինի, թմրամիջոցների, հակաբիոտիկների և մի շարք այլ դեղամիջոցների ազդեցության ժամանակ։
11. Կենդանու մարմնի ջերմաստիճանում խախտվում են սպերմատոգենեզի պրոցեսները։
12. Ցածր ջերմաստիճանի պայմաններում արու գամետները կարողանում են երկար ժամանակ պահպանել իրենց կենսական հատկությունները, ինչը հնարավորություն է տվել զարգացնել կենդանիների արհեստական բեղմնավորման տեխնոլոգիան։
13. Կանանց սեռական տրակտի բարենպաստ միջավայրում սպերմատոզոիդները պահպանում են իրենց բեղմնավորման կարողությունը 10-30 ժամ:
9. սպերմատոգենեզ
Այն իրականացվում է ամորձու ոլորված խողովակներում 4 փուլով.
1. վերարտադրության փուլ;
2. աճի փուլ;
3. հասունացման փուլ;
4. ձևավորման փուլ.
Բազմացման առաջին փուլում նկուղային թաղանթի վրա ընկած ցողունային բջիջները (քրոմոսոմների ամբողջական հավաքածուով) բազմիցս բաժանվում են միտոզով՝ առաջացնելով բազմաթիվ սերմնահեղուկներ։ Բաժանման յուրաքանչյուր փուլով դուստր բջիջներից մեկը մնում է այս ծայրահեղ շարքում որպես ցողունային բջիջ, մյուսը դուրս է մղվում հաջորդ շարքը և մտնում աճի փուլ:
Աճման փուլում սեռական բջիջները կոչվում են 1-ին կարգի սպերմատոցիտներ։ Նրանք աճում են և պատրաստվում են զարգացման երրորդ փուլին։ Այսպիսով, երկրորդ փուլը միաժամանակ ինտերֆազ է ապագա մեյոզից առաջ:
Հասունացման երրորդ փուլում սեռական բջիջները հաջորդաբար ենթարկվում են մեյոզի երկու բաժանման։ Միևնույն ժամանակ, 1-ին կարգի սպերմատոցիտներից ձևավորվում են 2-րդ կարգի սպերմատոցիտներ՝ կրկնապատկված քրոմոսոմների կես հավաքածուով։ Այս բջիջները կարճ ինտերֆազից հետո մտնում են մեյոզի երկրորդ բաժանում, որի արդյունքում առաջանում են սպերմատիդներ։ 2-րդ կարգի սպերմատոցիտները կազմում են սպերմատոգեն էպիթելի երրորդ շարքը։ Ինտերֆազի կարճ տեւողության պատճառով ոլորված խողովակների ողջ երկարությամբ 2-րդ կարգի սպերմատոցիտներ չեն հայտնաբերվում։ Սպերմատիդները խողովակների ամենափոքր բջիջներն են: Նրանք իրենց ներքին եզրերին կազմում են 2-3 բջիջների շարքեր։
Ձևավորման չորրորդ փուլում փոքր կլոր սերմնաբջիջները աստիճանաբար վերածվում են սպերմատոզոիդների, որոնք ունեն դրոշակի ձև: Այս գործընթացներն ապահովելու համար սպերմատիդները շփվում են տրոֆիկ Sertoli բջիջների հետ՝ ներթափանցելով իրենց ցիտոպլազմայի պրոցեսների միջև ընկած խորշերի մեջ։ Պատվիրված է միջուկի, շերտավոր համալիրի, ցենտրիոլների դասավորությունը։ Դիստալ ցենտրիոլից աճում է առանցքային թել, որից հետո ցիտոպլազմը տեղաշարժվում է պլազմոլեմայի հետ՝ ձևավորելով սերմնաբջիջի պոչը։ Շերտավոր բարդույթը գտնվում է միջուկի դիմաց և վերածվում է ակրոսոմի։ Միտոքոնդրիաները իջնում են բջջի մարմին՝ ձևավորվելով առանցքային պարուրաձև թելի շուրջ։ Ձևավորված սպերմատոզոիդների գլուխները դեռևս մնում են աջակից բջիջների խորշերում, իսկ դրանց պոչերը կախված են ոլորված խողովակի լույսի մեջ։
10. Ձվերի կառուցվածքը և դասակարգումը
Ձուն անշարժ, կլորաձև բջիջ է՝ դեղնուցի ներդիրների որոշակի պաշարով (ածխաջրային, սպիտակուցային և լիպիդային բնույթի սնուցիչներ)։ Հասուն ձվերում ցենտրոսոմներ չկան (դրանք կորչում են հասունացման փուլի վերջում)։
Կաթնասունների ձվերը, ի լրումն պլազմոլեմայի (օվոլեմման), որը հանդիսանում է առաջնային թաղանթ, ունեն նաև պաշտպանիչ և տրոֆիկ գործառույթներով երկրորդական թաղանթներ՝ փայլուն կամ թափանցիկ թաղանթ, որը բաղկացած է գլիկոզամինոգլիկաններից, սպիտակուցներից և մեկ շերտով ձևավորված շողացող պսակից։ պրիզմատիկ ֆոլիկուլային բջիջները սոսնձված են հիալուրոնաթթու:
Թռչունների մոտ երկրորդական թաղանթները թույլ են արտահայտված, սակայն զգալի զարգացած են երրորդական թաղանթները՝ սպիտակուցը, ենթափեղկերը, խեցիները և վերախորշը։ Նրանք գործում են որպես պաշտպանիչ և տրոֆիկ գոյացություններ հողային պայմաններում սաղմերի զարգացման ժամանակ։
Ձվաբջիջները դասակարգվում են ըստ քանակի և բաշխվածության դեղնուցի ցիտոպլազմայում.
1. Oligolecithal - փոքր դեղնուց ձու: Բնորոշ են բնակվող պարզունակ ակորդային կենդանիներին (նշտարակ)։ ջրային միջավայր, իսկ էգ կաթնասունները՝ կապված սաղմերի ներարգանդային զարգացման անցման հետ։
2. Մեզոլեցիտալ ձվաբջիջներ՝ դեղնուցի չափավոր կուտակումով։ Բնորոշ է ձկների և երկկենցաղների մեծամասնությանը:
3. Polylecital - բազմադեղնուց ձվերը բնորոշ են սողուններին և թռչուններին՝ կապված սաղմերի զարգացման ցամաքային պայմանների հետ։
Ձվի դասակարգումն ըստ դեղնուցի բաշխման.
1. Իզոլեցիտալ ձվեր, որոնցում դեղնուցի ներդիրները համեմատաբար հավասարաչափ բաշխված են ամբողջ ցիտոպլազմայում (նշտարակի և կաթնասունների օլիգոլեցիտալ ձվերը);
2. Տելոլեցիտալ ձու. Նրանց դեղնուցը տեղափոխվում է բջջի ստորին վեգետատիվ բևեռ, իսկ ազատ օրգանիլները և միջուկը տեղափոխվում են վերին կենդանական բևեռ (ձվերի միջուկային և տելոլեցիտալ տեսակներ ունեցող կենդանիների մոտ)։
11. Սաղմի զարգացման փուլերը
Սաղմնային զարգացումը փոխկապակցված փոխակերպումների շղթա է, որի արդյունքում միաբջիջ զիգոտից ձևավորվում է բազմաբջիջ օրգանիզմ, որն ունակ է գոյություն ունենալ արտաքին միջավայրում։ Էմբրիոգենեզում, որպես օնտոգենեզի մաս, արտացոլվում են նաև ֆիլոգենեզի գործընթացները։ Ֆիլոգենիան տեսակների պատմական զարգացումն է պարզից մինչև բարդ ձևեր: Օնտոգենեզը որոշակի օրգանիզմի անհատական զարգացումն է: Ըստ կենսագենետիկ օրենքի՝ օնտոգենեզը ֆիլոգենեզի կարճ ձև է, և, հետևաբար, կենդանիների տարբեր դասերի ներկայացուցիչներ ունեն սաղմի զարգացման ընդհանուր փուլեր.
1. Բեղմնավորում և զիգոտի ձևավորում;
2. Զիգոտի ճեղքում և բլաստուլայի ձևավորում;
3. Գաստրուլյացիա և երկու սաղմնային շերտերի առաջացում (էկտոդերմա և էնդոդերմա);
4. Էկտո- և էնդոդերմի տարբերակումը երրորդ սաղմնային շերտի՝ մեզոդերմի, առանցքային օրգանների (ակորդ, նյարդային խողովակ և առաջնային աղիքներ) և օրգանոգենեզի և հիստոգենեզի հետագա գործընթացների (օրգանների և հյուսվածքների զարգացում) առաջացմամբ:
Բեղմնավորումը ձվի և սերմնահեղուկի փոխադարձ յուրացման գործընթացն է, որի ժամանակ առաջանում է միաբջիջ օրգանիզմ՝ զիգոտը, որը միավորում է երկու ժառանգական տեղեկատվություն։
Զիգոտի ճեղքումը զիգոտի կրկնակի բաժանումն է միտոզով՝ առանց առաջացած բլաստոմերների աճի։ Այսպես է ձևավորվում ամենապարզ բազմաբջիջ օրգանիզմը՝ բլաստուլան։ Մենք առանձնացնում ենք.
Ամբողջական կամ հոլոբլաստիկ ջախջախիչ, որի ժամանակ ամբողջ զիգոտը մանրացված է բլաստոմերների (նշտարակ, երկկենցաղ, կաթնասուններ);
Անավարտ կամ մերոբլաստիկ, եթե զիգոտի միայն մի մասը (կենդանական բևեռ) ենթարկվում է ճեղքման (թռչուններ):
Ամբողջական ջախջախումը, իր հերթին, տեղի է ունենում.
Միատեսակ - իրենց համաժամանակյա բաժանմամբ ձևավորվում են համեմատաբար հավասար չափի բլաստոմերներ (նշտարակ);
Անհավասար - ասինխրոն բաժանմամբ տարբեր չափերի և ձևերի բլաստոմերների ձևավորմամբ (երկկենցաղներ, կաթնասուններ, թռչուններ):
Գաստրուլյացիան երկշերտ սաղմի ձևավորման փուլն է։ Նրա մակերեսային բջջային շերտը կոչվում է արտաքին սաղմնային շերտ՝ էկտոդերմա, իսկ խորը բջիջների շերտը՝ ներքին բողբոջային շերտ՝ էնդոդերմա։
Գաստրուլյացիայի տեսակները.
1. ինվագինացիա - բլաստուլայի հատակի բլաստոմերների ինվագինացիա տանիքի (նշտարակի) ուղղությամբ;
2. epiboly - աղտոտում իր եզրային գոտիների և հատակի բլաստուլայի տանիքի արագ բաժանվող փոքր բլաստոմերներով (երկկենցաղներ);
3. շերտազատում - բլաստոմերների շերտավորում և միգրացիա - բջիջների շարժում (թռչուններ, կաթնասուններ):
Սաղմնային շերտերի տարբերակումը հանգեցնում է տարբեր որակի բջիջների առաջացմանը, որոնք տալիս են տարբեր հյուսվածքների և օրգանների ռուդիմենտներ: Կենդանիների բոլոր դասերում առաջինը հայտնվում են առանցքային օրգանները՝ նյարդային խողովակը, նոտոկորդը, առաջնային աղիքը և երրորդ (միջին դիրքի) սաղմնային շերտը՝ մեզոդերմը։
12. Կաթնասունների սաղմնային զարգացման առանձնահատկությունները (տրոֆոբլաստների և պտղի թաղանթների ձևավորում)
Կաթնասունների էմբրիոգենեզի առանձնահատկությունները որոշվում են զարգացման ներարգանդային բնույթով, որի արդյունքում.
1. Ձուն դեղնուցի մեծ պաշարներ չի կուտակում (օլիգոլեցիտալ տեսակ)։
2. Բեղմնավորումը ներքին է:
3. Զիգոտի լրիվ անհավասար մասնատման փուլում տեղի է ունենում բլաստոմերների վաղ տարբերակում։ Նրանցից ոմանք ավելի արագ են բաժանվում, բնութագրվում են բաց գույնով և փոքր չափսերով, մյուսները մուգ գույնի են և մեծ չափերով, քանի որ այս բլաստոմերները ուշ են բաժանվում և ավելի քիչ են բաժանվում: Թեթև բլաստոմերներն աստիճանաբար պարուրում են դանդաղ բաժանվող մուգները, ինչի պատճառով առաջանում է գնդաձև բլաստուլա՝ առանց խոռոչի (մորուլա)։ Մորուլայում մուգ բլաստոմերները կազմում են դրա ներքին պարունակությունը բջիջների խիտ հանգույցի տեսքով, որոնք հետագայում օգտագործվում են սաղմի մարմինը կառուցելու համար. սա սաղմնաբլաստն է:
Թեթև բլաստոմերները գտնվում են սաղմի շուրջ մեկ շերտով։ Նրանց խնդիրն է կլանել արգանդի գեղձերի սեկրեցումը (արքայական ժելե), որպեսզի ապահովեն սաղմի սնուցման գործընթացները մինչև մոր օրգանիզմի հետ պլասենցային կապի ձևավորումը։ Հետեւաբար, նրանք ձեւավորում են տրոֆոբլաստ:
4. Բլաստուլայում արքայական ժելեի կուտակումը սաղմը դեպի վեր է մղում եւ նրան նմանեցնում թռչնի դիսկոբլաստուլայի։ Այժմ սաղմը ներկայացնում է բողբոջային վեզիկուլը կամ բլաստոցիստը: Արդյունքում, կաթնասունների հետագա զարգացման բոլոր գործընթացները կրկնում են թռչնի սաղմնածինին բնորոշ արդեն հայտնի ուղիները. գաստրուլյացիան իրականացվում է շերտազատման և միգրացիայի միջոցով; առանցքային օրգանների և մեզոդերմայի ձևավորումը տեղի է ունենում առաջնային շերտի և հանգույցի մասնակցությամբ, իսկ մարմնի մեկուսացումը և պտղի թաղանթների ձևավորումը՝ միջքաղաքային և ամնիոտիկ ծալքերը:
Բեռնախցիկի ծալքը ձևավորվում է բողբոջային վահանին սահմանակից գոտիներում բոլոր երեք սաղմնային շերտերի բջիջների ակտիվ վերարտադրության արդյունքում։ Բջիջների արագ աճը ստիպում է նրանց շարժվել դեպի ներս և թեքել տերևները։ Քանի որ միջքաղաքային ծալքը խորանում է, նրա տրամագիծը նվազում է, այն ավելի ու ավելի է առանձնանում և կլորացնում սաղմը, միաժամանակ ձևավորելով առաջնային աղիքն ու դեղնուցի պարկը դրա մեջ պարունակվող արքայական ժելեով էնդոդերմայից և ներքին օրգաններից:
Էկտոդերմայի ծայրամասային մասերը և մեզոդերմի պարիետալ թերթիկը կազմում են ամնիոտիկ շրջանաձև ծալք, որի եզրերն աստիճանաբար շարժվում են անջատված մարմնի վրայով և ամբողջովին փակվում նրա վրայով։ Ծալքի ներքին թիթեղների միաձուլումից առաջանում է ներքին ջրային թաղանթ՝ ամնիոն, որի խոռոչը լցված է ամնիոտիկ հեղուկով։ Ամնիոտիկ ծալքի արտաքին թաղանթների միաձուլումն ապահովում է պտղի ամենաարտաքին թաղանթի՝ քորիոնի (վիլոզային թաղանթի) ձևավորումը։
Առաջնային աղիքի փորային պատի պորտալարային ջրանցքով կույր ելքի պատճառով ձևավորվում է միջին թաղանթ՝ ալանտոիս, որում զարգանում է արյունատար անոթների համակարգ (անոթային թաղանթ)։
5. Արտաքին թաղանթ - քորիոնն ունի առանձնապես բարդ կառուցվածք և ձևավորում է բազմաթիվ ելուստներ վիլլի տեսքով, որոնց օգնությամբ սերտ հարաբերություններ են հաստատվում արգանդի լորձաթաղանթի հետ։ Վիլլիի բաղադրությունը ներառում է ալանտոիսի տարածքներ, որոնք միաձուլված են քորիոնի հետ արյան անոթների և տրոֆոբլաստների հետ, որոնց բջիջները հորմոններ են արտադրում հղիության բնականոն ընթացքը պահպանելու համար:
6. Ալանտոխորիոն վիլլիների և էնդոմետրիումի կառուցվածքների ամբողջությունը, որոնց հետ նրանք փոխազդում են, կաթնասունների մեջ կազմում են հատուկ սաղմնային օրգան՝ պլասենտան: Պլասենտան ապահովում է սաղմի սնուցումը, նրա գազափոխանակությունը, նյութափոխանակության արտադրանքի հեռացումը, ցանկացած էթիոլոգիայի անբարենպաստ գործոններից հուսալի պաշտպանություն և զարգացման հորմոնալ կարգավորում:
13. Պլասենտա (կառուցվածք, գործառույթներ, դասակարգումներ)
Պլասենտան ժամանակավոր օրգան է, որը ձևավորվում է կաթնասունների սաղմնային զարգացման ընթացքում։ Տարբերակել երեխայի և մայրական պլասենցայի միջև: Մանկական պլասենտան ձևավորվում է ալանտո-քորիոնիկ վիլլի հավաքածուով: Մայրը ներկայացված է արգանդի լորձաթաղանթի տարածքներով, որոնց հետ փոխազդում են այս վիլլիները:
Պլասենտան սաղմին ապահովում է սննդարար նյութերով (տրոֆիկ ֆունկցիա) և թթվածինով (շնչառական), սաղմի արյունը ազատում է ածխաթթու գազից և ավելորդ նյութափոխանակության արտադրանքներից (արտազատում), հորմոնների ձևավորում, որոնք ապահովում են հղիության բնականոն ընթացքը (էնդոկրին) և պլասենցային պատնեշի ձևավորում (պաշտպանիչ գործառույթ):
Պլասենցայի անատոմիական դասակարգումը հաշվի է առնում ալանտոխորիոնի մակերեսի վրա գտնվող վիլիների քանակը և գտնվելու վայրը:
1. Ցրված պլասենտան արտահայտված է խոզերի և ձիերի մոտ (կարճ, չճյուղավորված վիլլիները հավասարաչափ բաշխված են խորիոնի ամբողջ մակերեսով):
2. Բազմակի կամ կոթիլեդոնային պլասենտան բնորոշ է որոճողներին: Allantochorion villi-ները գտնվում են կղզիներում՝ կոթիլեդոններում:
3. Գոտիավոր պլասենտան մսակերների մոտ վիլլիների կուտակման գոտի է, որը գտնվում է պտղի միզապարկը շրջապատող լայն գոտու տեսքով:
4. Պրիմատների և կրծողների դիսկոիդային պլասենցայում խորիոնային վիլիզի գոտին ունի սկավառակի տեսք։
Պլասենցայի հյուսվածաբանական դասակարգումը հաշվի է առնում ալանտոխորիոն վիլիների փոխազդեցության աստիճանը արգանդի լորձաթաղանթի կառուցվածքների հետ: Ավելին, վիլլիների քանակի նվազումով դրանք ավելի ճյուղավորված են իրենց ձևով և ավելի խորը թափանցում արգանդի լորձաթաղանթի մեջ՝ կրճատելով սննդանյութերի շարժման ճանապարհը։
1. Epitheliochorial placenta-ն բնորոշ է խոզերին, ձիերին։ Chorionic villi թափանցում է արգանդի խցուկներ, առանց ոչնչացնելու էպիթելի շերտը: Ծննդաբերության ժամանակ վիլլիները հեշտությամբ դուրս են գալիս արգանդի գեղձերից՝ սովորաբար առանց արյունահոսության, ուստի պլասենցայի այս տեսակը կոչվում է նաև կիսապլասենտա։
2. Դեզմոխորիալ պլասենտան արտահայտված է որոճողների մոտ։ Ալանտո-քորիոնիկ վիլլիները ներթափանցում են էնդոմետրիումի լամինա պրոպրիա, դրա խտացումների, կարունկների հատվածում:
3. Էնդոթելիոխորիալ պլասենտան բնորոշ է մսակեր կենդանիներին։ Մանկական պլասենցայի վիլլիները շփվում են արյան անոթների էնդոթելիի հետ:
4. Հեմոխորիալ պլասենտան հանդիպում է պրիմատների մոտ։ Քորիոնիկ վիլլիները սուզվում են արյունով լցված բացվածքների մեջ և ողողվում մայրական արյունով։ Այնուամենայնիվ, մոր արյունը չի խառնվում պտղի արյան հետ:
14. Մորֆոլոգիական դասակարգումը և էպիթելի հիմնական տեսակների համառոտ նկարագրությունը
Էպիթելային հյուսվածքների մորֆոլոգիական դասակարգումը հիմնված է երկու հատկանիշների վրա.
1. էպիթելային բջիջների շերտերի քանակը;
2. բջջի ձևը. Այս դեպքում շերտավորված էպիթելի տեսակների մեջ հաշվի է առնվում միայն մակերեսային (տարածքային) շերտի էպիթելիոցիտների ձևը։
Միաշերտ էպիթելի, ի լրումն, կարող է կառուցվել նույն ձևի և բարձրության բջիջներից, այնուհետև դրանց միջուկները գտնվում են նույն մակարդակի վրա՝ մեկ շարքով էպիթելի և զգալիորեն տարբեր էպիթելիոցիտներից:
Նման դեպքերում ցածր բջիջներում միջուկները կկազմեն ստորին շարքը, միջին չափի էպիթելային բջիջներում՝ հաջորդը, որը գտնվում է առաջինից վերև, իսկ ամենաբարձրում՝ ևս մեկ կամ երկու շարք միջուկներ, ինչը, ի վերջո, թարգմանում է. միաշերտ հյուսվածքը վերածվում է կեղծ բազմաշերտ ձևի՝ բազմաշերտ էպիթելիի:
Հաշվի առնելով վերը նշվածը, էպիթելի մորֆոլոգիական դասակարգումը կարող է ներկայացվել հետևյալ կերպ.
Էպիթելիա
Միաշերտ Բազմաշերտ
Մեկ տող Բազմաշար հարթ. Անցումային խորանարդ
Flat Prismatic keratinizing
Խորանարդ թարթիչավոր չկերատինացնող
Prismatic- (ciliated) Prismatic
Ցանկացած տեսակի միաշերտ էպիթելիում նրա յուրաքանչյուր բջիջ կապ ունի նկուղային թաղանթի հետ։ Ցողունային բջիջները գտնվում են խճանկարային ձևով ծածկույթների միջև:
Շերտավորված էպիթելում մենք առանձնացնում ենք էպիթելիոցիտների երեք գոտի, որոնք տարբերվում են ձևով և տարբերակման աստիճանով: Պրիզմատիկ կամ բարձր խորանարդ բջիջների միայն ամենացածր շերտը կապված է նկուղային թաղանթի հետ: Այն կոչվում է բազալ և բաղկացած է ցողունից՝ բազմիցս բաժանվող էպիթելիոցիտներից։ Հաջորդ, միջանկյալ գոտին ներկայացված է տարբեր ձևերի տարբերվող (հասունացող) բջիջներով, որոնք կարող են ընկած լինել մեկ կամ մի քանի շարքերում։ Մակերեւույթում կան որոշակի ձևի և հատկությունների հասուն տարբերակված էպիթելիոցիտներ։ Շերտավորված էպիթելը ապահովում է պաշտպանիչ գործառույթներ:
Միաշերտ տափակ էպիթելը ձևավորվում է անկանոն եզրագծերով և մեծ մակերեսով հարթեցված բջիջներով։ ծածկում է շիճուկային թաղանթները (մեզոթելիում); կազմում է թոքերի անոթային լորձաթաղանթը (էնդոթելիում) և ալվեոլները (շնչառական էպիթելը)։
Միաշերտ խորանարդ էպիթելը կառուցված է էպիթելային բջիջներից, որոնք ունեն մոտավորապես նույն հիմքի լայնությունը և բարձրությունը: Միջուկը կլորացված է, բնութագրվում է կենտրոնական դիրքով։ Ձևավորում է գեղձերի արտազատիչ հատվածները, միզուղիների երիկամային խողովակների (նեֆրոնների) պատերը։
Միաշերտ պրիզմատիկ էպիթելը կազմում է էկզոկրին գեղձերի՝ արգանդի գեղձերի արտազատվող խողովակների պատերը, ծածկում է աղիքային տիպի ստամոքսի, բարակ և հաստ աղիքի լորձաթաղանթը։ Բջիջները բնութագրվում են բարձր բարձրությամբ, նեղ հիմքով և դեպի բազալ բևեռ տեղափոխված միջուկի երկայնական օվալաձև ձևով։ Աղիքային էպիթելը սահմանափակվում է էնտերոցիտների գագաթային բևեռներում միկրովիլիներով:
Միաշերտ բազմաշերտ պրիզմատիկ թարթիչավոր (ciliated) էպիթելը ծածկում է հիմնականում շնչուղիների լորձաթաղանթը։ Ամենացածր սեպաձև բջիջները (բազալ) անընդհատ բաժանվում են, միջինները աճում են բարձրության վրա, դեռևս չեն հասնում ազատ մակերեսին, իսկ բարձրերը հասուն էպիթելի բջիջների հիմնական տեսակն են, որոնք գագաթային բևեռներում կրում են մինչև 300 թարթիչներ։ , որոնք կծկվելով՝ տեղափոխում են լորձը կլանված օտար մասնիկներով հազի համար։ Լորձը արտադրվում է թարթիչավոր գավաթային բջիջներով:
Շերտավորված տափակ ոչ կերատինացված էպիթելը ծածկում է աչքերի կոնյուկտիվան և եղջերաթաղանթը, մարսողական խողովակի սկզբնական հատվածները, վերարտադրության և միզուղիների արտազատման օրգանների անցումային գոտիները:
Շերտավորված տափակ կերատինացված էպիթելը բաղկացած է աստիճանաբար կերատինացնող և շերտազատվող բջիջների 5 շերտից (կերատինոցիտներ)՝ բազալ, փշոտ բջիջների շերտ, հատիկավոր, փայլուն, եղջյուրավոր։ Այն կազմում է մաշկի էպիդերմիսը, ծածկում արտաքին սեռական օրգանները, կաթնագեղձերի կաթնագեղձերի լորձաթաղանթը և բերանի խոռոչի մեխանիկական պապիլները։
Շերտավորված անցումային էպիթելը գծում է միզուղիների լորձաթաղանթները: Ներծծված գոտու բջիջները մեծ են, երկայնական օվալաձև են, արտազատում են լորձ, ունեն լավ զարգացած գլիկոկալիքս պլազմոլեմայում՝ կանխելու համար մեզից նյութերի հակադարձ կլանումը։
Շերտավորված պրիզմատիկ էպիթելն արտահայտվում է պարիետալ թքագեղձերի հիմնական ծորանների բերաններում, տղամարդկանց մոտ՝ միզասեռական ջրանցքի կոնքի հատվածի լորձաթաղանթում և ամորձիների կցորդների ջրանցքներում, կանանց մոտ՝ բլթակային խողովակներում։ կաթնագեղձերը՝ երկրորդական և երրորդական ձվարանների ֆոլիկուլներում։
Բազմաշերտ խորանարդը կազմում է մաշկի ճարպագեղձերի արտազատիչ հատվածները, իսկ արական սեռի մոտ՝ ամորձիների ոլորված խողովակների սպերմատոգեն էպիթելը:
15. Արյան ընդհանուր բնութագրերը որպես մարմնի ներքին միջավայրի հյուսվածք
Արյունը պատկանում է հենակետային-տրոֆիկ խմբի հյուսվածքներին։ Ցանցային և չամրացված շարակցական հյուսվածքների հետ որոշիչ դեր է խաղում մարմնի ներքին միջավայրի ձևավորման գործում։ Այն ունի հեղուկ հետևողականություն և իրենից ներկայացնում է համակարգ, որը բաղկացած է երկու բաղադրիչներից՝ միջբջջային նյութից (պլազմա) և դրանում կախված բջիջներից՝ ձևավորված տարրերից՝ էրիթրոցիտներ, լեյկոցիտներ և թրոմբոցիտներ (կաթնասունների արյան թրոմբոցիտներ):
Պլազման կազմում է արյան զանգվածի մոտ 60%-ը և պարունակում է 90-93% ջուր և 7-10% պինդ նյութեր։ Դրա մոտ 7%-ը բաժին է ընկնում սպիտակուցներին (4%-ը՝ ալբումիններ, 2,8%-ը՝ գլոբուլիններ և 0,4%-ը՝ ֆիբրինոգեն), 1%-ը՝ հանքային նյութերին, նույն տոկոսը մնում է ածխաջրերին։
Արյան պլազմայի սպիտակուցների գործառույթները.
Ալբոմիններ. - թթու-բազային հավասարակշռության կարգավորում;
Տրանսպորտ;
Օսմոտիկ ճնշման որոշակի մակարդակի պահպանում.
Գլոբուլինները իմունային սպիտակուցներ են (հակամարմիններ), որոնք կատարում են պաշտպանիչ գործառույթ և մի շարք ֆերմենտային համակարգեր:
Ֆիբրինոգեն - մասնակցում է արյան մակարդման գործընթացներին:
Արյան pH-ը 7,36 է և բավականին կայուն է այս մակարդակում մի շարք բուֆերային համակարգերի կողմից:
Արյան հիմնական գործառույթները.
1. Անընդհատ շրջանառվելով արյունատար անոթներով՝ իրականացնում է թթվածնի փոխանցումը թոքերից հյուսվածքներ, և. ածխաթթու գազհյուսվածքներից մինչև թոքեր (գազի փոխանակման գործառույթ); Մարսողական համակարգում ներծծված սննդանյութերը մատակարարում է մարմնի բոլոր օրգաններին, իսկ նյութափոխանակության արտադրանքները արտազատող օրգաններին (տրոֆիկ); տեղափոխում է հորմոններ, ֆերմենտներ և այլ կենսաբանական ակտիվ նյութեր նրանց ակտիվ ազդեցության վայրեր:
Արյան ֆունկցիոնալ գործառույթների այս բոլոր ասպեկտները կարող են կրճատվել մեկ ընդհանուր տրանսպորտային և տրոֆիկ ֆունկցիայի:
2. Հոմեոստատիկ - մարմնի ներքին միջավայրի կայունության պահպանում (օպտիմալ պայմաններ է ստեղծում նյութափոխանակության ռեակցիաների համար);
3. Պաշտպանիչ - ապահովում է բջջային և հումորալ իմունիտետ, ոչ սպեցիֆիկ պաշտպանության տարբեր ձևեր, հատկապես օտար մասնիկների ֆագոցիտոզ, արյան մակարդման պրոցեսներ։
4. Կարգավորող ֆունկցիա՝ կապված մարմնի մշտական ջերմաստիճանի պահպանման և մի շարք այլ պրոցեսների հետ, որոնք ապահովվում են հորմոններով և այլ կենսաբանական ակտիվ նյութերով:
Թրոմբոցիտներ - կաթնասունների մոտ արյան մակարդման գործընթացներում ներգրավված են ոչ միջուկային բջիջներ, 3-5 մկմ չափերով։
Լեյկոցիտները բաժանվում են գրանուլոցիտների (բազոֆիլներ, նեյտրոֆիլներ և էոզինոֆիլներ) և ագրանուլոցիտների (մոնոցիտներ և լիմֆոցիտներ): Նրանք կատարում են տարբեր պաշտպանիչ գործառույթներ:
Կաթնասունների էրիթրոցիտները ոչ միջուկային բջիջներ են, դրանք 6-8 մկմ միջին տրամագծով երկգոգավոր սկավառակների տեսքով են։
Արյան պլազմայի մի մասը միկրոանոթային անոթների միջոցով անընդհատ մտնում է օրգանների հյուսվածքներ և դառնում հյուսվածքային հեղուկ: Տալով սննդանյութեր, ընկալելով նյութափոխանակության արգասիքները, հարստանալով արյունաստեղծ օրգաններում լիմֆոցիտներով, վերջիններս ավիշի տեսքով մտնում են ավշային համակարգի անոթներ և վերադառնում արյան հոսք։
Արյան մեջ ձևավորված տարրերը որոշակի քանակական հարաբերակցության մեջ են և կազմում են նրա հեմոգրամը։
Ձևավորված տարրերի քանակը հաշվարկվում է 1 մկլ արյան կամ լիտրի մեջ.
Էրիտրոցիտներ - 5-10 միլիոն մեկ μl (x 1012 մեկ լ);
Լեյկոցիտներ - 4,5-14 հազար մեկ μl (x109 մեկ լ);
Արյան թրոմբոցիտներ - 250-350 հազար մեկ մկլ (x109 մեկ լ):
16. Գրանուլոցիտների կառուցվածքը և ֆունկցիոնալ նշանակությունը
Ողնաշարավորների լեյկոցիտները միջուկային բջիջներ են, որոնք ունակ են ակտիվ շարժվել մարմնի հյուսվածքներում: Դասակարգումը հիմնված է հաշվի առնելով դրանց ցիտոպլազմայի կառուցվածքային առանձնահատկությունները:
Լեյկոցիտները, որոնց ցիտոպլազմը պարունակում է հատուկ հատիկավորություն, կոչվում են հատիկավոր կամ գրանուլոցիտներ։ Հասուն հատիկավոր լեյկոցիտներն ունեն հատվածավորված միջուկ՝ հատվածավորված բջիջներ, երիտասարդների մոտ այն չհատված է: Հետևաբար, ընդունված է դրանք բաժանել երիտասարդ ձևերի (լոբի ձևավորված միջուկ), դանակ-միջուկային (կոր ձողաձև միջուկ) և սեգմենտավորված - լիովին տարբերակված լեյկոցիտների, որոնց միջուկը պարունակում է 2-ից 5-7 հատված: Համաձայն ցիտոպլազմային հատիկավորության ներկման տարբերության՝ գրանուլոցիտների խմբում առանձնանում են 3 տեսակի բջիջներ.
Բազոֆիլներ - հատիկավորությունը ներկված է հիմնական ներկերով մանուշակագույնով;
Էոզինոֆիլներ - հատիկավորությունը ներկված է թթվային ներկերով կարմիրի տարբեր երանգներով;
Նեյտրոֆիլներ - հատիկավորությունը ներկված է ինչպես թթվային, այնպես էլ հիմնային ներկերով վարդագույն-մանուշակագույն գույնով:
Նեյտրոֆիլները փոքր բջիջներ են (9-12 մկմ), որոնց ցիտոպլազմը պարունակում է 2 տեսակի հատիկներ՝ առաջնային (բազոֆիլ), որոնք լիզոսոմներ են և երկրորդային օքսիֆիլ (պարունակում են կատիոնային սպիտակուցներ և ալկալային ֆոսֆատազ)։ Նեյտրոֆիլները բնութագրվում են ամենալավ (փոշու նման) հատիկավորությամբ և ամենահատված միջուկով։ Նրանք միկրոֆագներ են և իրականացնում են ցանկացած բնույթի մանր օտար մասնիկների ֆագոցիտային ֆունկցիա, հակագեն-հակամարմին կոմպլեքսների օգտագործում։ Բացի այդ, արտազատվում են նյութեր, որոնք խթանում են վնասված հյուսվածքների վերականգնումը։
Էոզինոֆիլները հաճախ պարունակում են երկու հատվածի միջուկ և մեծ օքսիֆիլային հատիկներ ցիտոպլազմայում: Նրանց տրամագիծը 12-18 միկրոն է։ Հատիկները պարունակում են հիդրոլիտիկ ֆերմենտներ (գործառույթով միկրոֆագներ): Նրանք ցույց են տալիս հակահիստամինային ռեակտիվություն, խթանում են շարակցական հյուսվածքի մակրոֆագների ֆագոցիտային ակտիվությունը և դրանցում լիզոսոմների ձևավորումը, օգտագործում են հակագեն-հակամարմին համալիրներ: Բայց նրանց հիմնական խնդիրն է չեզոքացնել թունավոր նյութերը, ուստի էոզինոֆիլների թիվը կտրուկ աճում է հելմինթիկ ներխուժումներով:
Բազոֆիլները՝ 12-16 միկրոն չափերով, պարունակում են միջին չափի բազոֆիլային հատիկներ, որոնք ներառում են հեպարին (կանխում է արյան մակարդումը) և հիստամին (կարգավորում է անոթների և հյուսվածքների թափանցելիությունը)։ Նրանք նաև մասնակցում են ալերգիկ ռեակցիաների զարգացմանը:
Լեյկոցիտների առանձին տեսակների տոկոսային հարաբերակցությունը կոչվում է լեյկոցիտային բանաձև կամ լեյկոգրամ: Գրանուլոցիտների համար այն ունի հետևյալ տեսքը.
Նեյտրոֆիլներ - 25-40% - խոզերի և որոճողների մեջ; 50-70% - ձիերի և մսակերների մեջ;
Էոզինոֆիլներ՝ 2-4%, որոճողների մոտ՝ 6-8%;
Բազոֆիլներ - 0,1-2%:
17. Ագրանուլոցիտների կառուցվածքը և ֆունկցիոնալ նշանակությունը
Ոչ հատիկավոր լեյկոցիտները (ագրանուլոցիտները) բնութագրվում են ցիտոպլազմայում հատուկ հատիկավորության բացակայությամբ և խոշոր ոչ հատվածավորված միջուկներում։ Ագրանուլոցիտների խմբում առանձնանում են բջիջների 2 տեսակ՝ լիմֆոցիտներ և մոնոցիտներ։
Լիմֆոցիտներին բնորոշ է միջուկի գերակշռող կլոր ձևը՝ կոմպակտ քրոմատինով։ Փոքր լիմֆոցիտներում միջուկը զբաղեցնում է գրեթե ամբողջ բջիջը (դրա տրամագիծը 4,5-6 մկմ է), միջին չափի լիմֆոցիտներում ցիտոպլազմայի եզրն ավելի լայն է, և դրանց տրամագիծը մեծանում է մինչև 7-10 մկմ։ Ծայրամասային արյան մեջ մեծ լիմֆոցիտները (10-13 միկրոն) չափազանց հազվադեպ են: Լիմֆոցիտների ցիտոպլազմը ներկվում է բազոֆիլ կերպով՝ կապույտի տարբեր երանգներով։
Լիմֆոցիտներն ապահովում են բջջային և հումորալ իմունիտետի ձևավորում։ Դրանք դասակարգվում են T- և B-լիմֆոցիտների:
T-լիմֆոցիտները (ուրմուսից կախված) ենթարկվում են առաջնային հակագենից անկախ տարբերակման տիմուսում: Իմունային համակարգի ծայրամասային օրգաններում, անտիգենների հետ շփվելուց հետո, դրանք վերածվում են պայթյունի ձևերի, բազմապատկվում և այժմ ենթարկվում են երկրորդական հակագենակախյալ տարբերակման, ինչի արդյունքում առաջանում են T բջիջների էֆեկտորային տեսակներ.
T-մարդասպաններ, որոնք ոչնչացնում են օտար և իրենց սեփական բջիջները թերի ֆենոկոպներով (բջջային իմունիտետ);
T-helpers - խթանում է B-լիմֆոցիտների վերափոխումը պլազմային բջիջների;
T-suppressors, որոնք ճնշում են B-լիմֆոցիտների ակտիվությունը;
Հիշողության T-լիմֆոցիտներ (երկարակյաց բջիջներ), որոնք պահպանում են տեղեկատվություն անտիգենների մասին:
B-լիմֆոցիտներ (բուրսոկախված): Թռչունների մոտ դրանք հիմնականում տարբերվում են Ֆաբրիցիուսի բուրսայում, իսկ կաթնասունների մոտ՝ կարմիր ոսկրածուծում։ Երկրորդային տարբերակման ժամանակ դրանք վերածվում են պլազմային բջիջների, որոնք արտադրում են մեծ քանակությամբ հակամարմիններ, որոնք մտնում են արյուն և այլ մարմնի հեղուկներ, ինչը ապահովում է անտիգենների չեզոքացումը և հումորալ իմունիտետի ձևավորումը։
Մոնոցիտները արյան ամենամեծ բջիջներն են (18-25 մկմ): Միջուկը երբեմն լոբի տեսք ունի, բայց ավելի հաճախ՝ անկանոն։ Ցիտոպլազմը զգալիորեն արտահայտված է, նրա մասնաբաժինը կարող է հասնել բջջի ծավալի կեսին, այն ներկվում է բազոֆիլորեն՝ ծխագույն կապույտ գույնով։ Այն ունի լավ զարգացած լիզոսոմներ։ Արյան մեջ շրջանառվող մոնոցիտները հյուսվածքների և օրգանների մակրոֆագների պրեկուրսորներն են, որոնք մարմնում կազմում են պաշտպանիչ մակրոֆագային համակարգ՝ մոնոմիջուկային ֆագոցիտների համակարգը (MPS): Անոթային արյան մեջ կարճատև մնալուց հետո (12-36 ժամ) մոնոցիտները մազանոթների և վենուլների էնդոթելի միջոցով գաղթում են հյուսվածքներ և վերածվում ֆիքսված և ազատ մակրոֆագների։
Մակրոֆագները հիմնականում օգտագործում են մեռնող և վնասված բջջային և հյուսվածքային տարրեր: Բայց նրանք ավելի պատասխանատու դեր են խաղում իմունային ռեակցիաներում.
Նրանք փոխակերպում են անտիգենները մոլեկուլային ձևի և ներկայացնում դրանք լիմֆոցիտների (հակագին ներկայացնող ֆունկցիա):
Նրանք արտադրում են ցիտոկիններ՝ խթանելու T և B բջիջները:
Օգտագործեք հակագենների համալիրներ հակամարմիններով:
Լեյկոգրամում ագրանուլոցիտների տոկոսը.
Մոնոցիտներ - 1-8%;
Լիմֆոցիտները՝ գիշատիչ կենդանիների և ձիերի մոտ՝ 20-40%, խոզերի մոտ՝ 45-56%, խոշոր եղջերավորների մոտ՝ 45-65%:
18. Չամրացված շարակցական հյուսվածքի մորֆոֆունկցիոնալ բնութագրերը
Չամրացված շարակցական հյուսվածքը առկա է բոլոր օրգաններում և հյուսվածքներում՝ հիմք հանդիսանալով էպիթելի, գեղձերի տեղադրման համար՝ օրգանների ֆունկցիոնալ կառուցվածքները միացնելով մեկ համակարգի մեջ: Ուղեկցում է արյունատար անոթներին և նյարդերին։ Այն կատարում է ձևավորող, աջակցող, պաշտպանիչ և տրոֆիկ գործառույթներ: Հյուսվածքը բաղկացած է բջիջներից և միջբջջային նյութից։ Սա պոլիդիֆերենցիալ գործվածք է, քանի որ. նրա բջիջները ստացվել են տարբեր ցողունային բջիջներից:
Նմանատիպ փաստաթղթեր
Հյուսվածքաբանությունը կենդանական օրգանիզմների և մարդու մարմնի հյուսվածքների զարգացման, կառուցվածքի, կենսագործունեության և վերածնման ուսումնասիրությունն է: Նրա հետազոտության մեթոդները, զարգացման փուլերը, առաջադրանքները: Համեմատական սաղմնաբանության հիմունքներ, գիտություն մարդու սաղմի զարգացման և կառուցվածքի մասին։
վերացական, ավելացվել է 12/01/2011 թ
Հյուսվածքաբանություն - գիտություն կենդանիների օրգանիզմների հյուսվածքների կառուցվածքի, զարգացման և կենսագործունեության և հյուսվածքների կազմակերպման ընդհանուր ձևերի մասին. բջջաբանության և սաղմնաբանության հայեցակարգը: Հյուսվածքաբանական հետազոտության հիմնական մեթոդները; հյուսվածքաբանական պատրաստուկի պատրաստում.
շնորհանդես, ավելացվել է 03/23/2013
Հյուսվածքաբանության պատմություն - կենսաբանության ճյուղ, որն ուսումնասիրում է կենդանի օրգանիզմների հյուսվածքների կառուցվածքը։ Հետազոտության մեթոդներ հյուսվածաբանության մեջ, հյուսվածքաբանական պատրաստուկի պատրաստում։ Հյուսվածքների հյուսվածաբանություն - բջիջների և ոչ բջջային կառուցվածքների ֆիլոգենետիկ ձևավորված համակարգ:
վերացական, ավելացվել է 01/07/2012 թ
Հիստոլոգիայի հիմնական դրույթները, որոնք ուսումնասիրում են բջիջների համակարգը, ոչ բջջային կառուցվածքները, որոնք ունեն ընդհանուր կառուցվածք և ուղղված են որոշակի գործառույթների իրականացմանը: Էպիթելի, արյան, ավշի, միացնող, մկանային, նյարդային հյուսվածքի կառուցվածքի, ֆունկցիաների վերլուծություն։
վերացական, ավելացվել է 23.03.2010թ
Մարդու տարբեր հյուսվածքների տեսակների և գործառույթների ուսումնասիրություն: Հյուսվածքաբանության գիտության խնդիրները, որն ուսումնասիրում է կենդանի օրգանիզմների հյուսվածքների կառուցվածքը։ Էպիթելի, նյարդային, մկանային հյուսվածքի և ներքին միջավայրի հյուսվածքների կառուցվածքի առանձնահատկությունները (միացնող, կմախքային և հեղուկ):
ներկայացում, ավելացվել է 11/08/2013 թ
Հյուսվածքաբանության ուսումնասիրության հիմնական առարկան. Հյուսվածքաբանական վերլուծության հիմնական փուլերը, դրա ուսումնասիրության օբյեկտները. Լույսի և էլեկտրոնային մանրադիտակի համար հյուսվածաբանական պատրաստուկի պատրաստման գործընթացը: Լյումինեսցենտային (լյումինեսցենտ) մանրադիտակ, մեթոդի էությունը.
կուրսային աշխատանք, ավելացվել է 01.12.2015թ
Կենդանի բջիջների հիմնական տեսակները և դրանց կառուցվածքի առանձնահատկությունները. Էուկարիոտ և պրոկարիոտ բջիջների կառուցվածքի ընդհանուր պլանը. Բույսերի և սնկային բջիջների կառուցվածքի առանձնահատկությունները. Բույսերի, կենդանիների, սնկերի և բակտերիաների բջիջների կառուցվածքի համեմատական աղյուսակ.
վերացական, ավելացվել է 12.01.2016թ
Լույսի մանրադիտակի համար հյուսվածքաբանական պատրաստուկների պատրաստման տեխնիկան, այս գործընթացի հիմնական փուլերը և դրա իրականացման պայմաններին ներկայացվող պահանջները: Հետազոտության մեթոդներ հյուսվածաբանության և բջջաբանության մեջ: Հեմատոքսիլին-էոզինի պատրաստուկների ներկման մոտավոր սխեման:
թեստ, ավելացվել է 10/08/2013
Սպերմատոգենեզի, միտոտիկ բջիջների բաժանման բնութագրերը՝ ըստ մեյոզի տեսակի. Բջջային տարբերակման փուլերի ուսումնասիրությունը, որոնք միասին կազմում են սերմնաբուծական էպիթելը։ Արական սեռական օրգանների և դրանց գեղձերի կառուցվածքի, շագանակագեղձի ֆունկցիաների ուսումնասիրությունը։
վերացական, ավելացվել է 12/05/2011 թ
Հյուսվածքաբանության ծննդյան պատմությունը որպես գիտություն. Հյուսվածքաբանական պատրաստուկներ և դրանց ուսումնասիրության մեթոդներ. Հյուսվածքաբանական պատրաստուկների պատրաստման փուլերի բնութագրումը՝ ամրացում, լարեր, լցնում, կտրում, ներկում և հատվածավորում։ Մարդու հյուսվածքների տիպաբանություն.

 Հյուսվածքը բջիջների և դրանց ածանցյալների (մանրաթելեր, ամորֆ նյութ, սինցիտիա, սիմպլաստներ) համակարգ է, որն առաջացել է զարգացման գործընթացում և բնութագրվում է ընդհանուր մորֆոֆիզիոլոգիական հատկություններով։ Syncytium կոչվում է ցանցային կառուցվածք, որը բաղկացած է բջիջներից, որոնց գործընթացները սերտորեն փոխկապակցված են: Սիմպլաստը մի կառույց է, որը բաղկացած է բազմաթիվ բջիջներից, որոնք միաձուլվել են միմյանց հետ (այսպես է կառուցվում գծավոր մկանային հյուսվածքը)։
Հյուսվածքը բջիջների և դրանց ածանցյալների (մանրաթելեր, ամորֆ նյութ, սինցիտիա, սիմպլաստներ) համակարգ է, որն առաջացել է զարգացման գործընթացում և բնութագրվում է ընդհանուր մորֆոֆիզիոլոգիական հատկություններով։ Syncytium կոչվում է ցանցային կառուցվածք, որը բաղկացած է բջիջներից, որոնց գործընթացները սերտորեն փոխկապակցված են: Սիմպլաստը մի կառույց է, որը բաղկացած է բազմաթիվ բջիջներից, որոնք միաձուլվել են միմյանց հետ (այսպես է կառուցվում գծավոր մկանային հյուսվածքը)։
 Հյուսվածքների բոլոր տեսակները միավորված են չորս հիմնական խմբերում՝ 1) էպիթելային, 2) հենարանային-տրոֆիկ, 3) մկանային, 4) նյարդային հյուսվածքի։
Հյուսվածքների բոլոր տեսակները միավորված են չորս հիմնական խմբերում՝ 1) էպիթելային, 2) հենարանային-տրոֆիկ, 3) մկանային, 4) նյարդային հյուսվածքի։
 Էպիթելային հյուսվածքներ Օրգանիզմի և շրջակա միջավայրի միջև սահմանի վրա ամենուր, այն առանձնացնելով շրջակա միջավայրից - այն ծածկում է մարմինը մակերեսից շարունակական շերտով և գծում ներքին օրգանները. կա էպիթելային հյուսվածք:
Էպիթելային հյուսվածքներ Օրգանիզմի և շրջակա միջավայրի միջև սահմանի վրա ամենուր, այն առանձնացնելով շրջակա միջավայրից - այն ծածկում է մարմինը մակերեսից շարունակական շերտով և գծում ներքին օրգանները. կա էպիթելային հյուսվածք:
 Բոլոր էպիթելիաները կառուցված են էպիթելային բջիջներից՝ էպիթելիոցիտներից: Էպիթելիոցիտները միանում են միմյանց դեսմոսոմների, փակող ժապավենների, սոսնձող ժապավենների օգնությամբ՝ կազմելով բջջային շերտ։ Էպիթելային շերտերը կցվում են նկուղային թաղանթին, իսկ դրա միջոցով՝ էպիթելը սնուցող շարակցական հյուսվածքին։
Բոլոր էպիթելիաները կառուցված են էպիթելային բջիջներից՝ էպիթելիոցիտներից: Էպիթելիոցիտները միանում են միմյանց դեսմոսոմների, փակող ժապավենների, սոսնձող ժապավենների օգնությամբ՝ կազմելով բջջային շերտ։ Էպիթելային շերտերը կցվում են նկուղային թաղանթին, իսկ դրա միջոցով՝ էպիթելը սնուցող շարակցական հյուսվածքին։
 Նկուղային թաղանթը բաղկացած է ամորֆ նյութից և ֆիբրիլային կառուցվածքներից:Նկուղային թաղանթի գործառույթներն են մակրոմոլեկուլային միացությունների տեղափոխումը և էպիթելի բջիջների առաձգական հիմքի ստեղծումը: Հյուսվածքը չունի արյունատար անոթներ, չկա ոչ բջջային ձևեր: էպիթելի բջիջները սնվում են շարակցական հյուսվածքից եկող հյուսվածքային հեղուկով:
Նկուղային թաղանթը բաղկացած է ամորֆ նյութից և ֆիբրիլային կառուցվածքներից:Նկուղային թաղանթի գործառույթներն են մակրոմոլեկուլային միացությունների տեղափոխումը և էպիթելի բջիջների առաձգական հիմքի ստեղծումը: Հյուսվածքը չունի արյունատար անոթներ, չկա ոչ բջջային ձևեր: էպիթելի բջիջները սնվում են շարակցական հյուսվածքից եկող հյուսվածքային հեղուկով:
 Կախված գտնվելու վայրից և կատարվող ֆունկցիայից՝ առանձնանում են էպիթելի երկու տեսակ՝ ծածկված և գեղձային։
Կախված գտնվելու վայրից և կատարվող ֆունկցիայից՝ առանձնանում են էպիթելի երկու տեսակ՝ ծածկված և գեղձային։
 Բջիջների գտնվելու վայրի բնույթով ծածկված էպիթելիումը բաժանվում է. իսկ մնացած բոլորը գտնվում են հիմքում ընկած էպիթելի բջիջների վրա):
Բջիջների գտնվելու վայրի բնույթով ծածկված էպիթելիումը բաժանվում է. իսկ մնացած բոլորը գտնվում են հիմքում ընկած էպիթելի բջիջների վրա):
 Միաշերտ էպիթելի մի շարք (բջիջների և միջուկների ազատ ծայրերը գտնվում են նույն մակարդակի վրա) բազմաշարք (բոլոր բջիջները ընկած են նկուղային թաղանթի վրա, բայց միջուկները գտնվում են դրանից տարբեր բարձրության վրա, ինչը հանգեցնում է բազմաշարժ էֆեկտի: )
Միաշերտ էպիթելի մի շարք (բջիջների և միջուկների ազատ ծայրերը գտնվում են նույն մակարդակի վրա) բազմաշարք (բոլոր բջիջները ընկած են նկուղային թաղանթի վրա, բայց միջուկները գտնվում են դրանից տարբեր բարձրության վրա, ինչը հանգեցնում է բազմաշարժ էֆեկտի: )

 Integumentary epithelium (սխեմա ըստ Ալեքսանդրովսկայայի). B - խորանարդ; B - գլանաձեւ (սյունակ); G - բազմաշերտ գլանաձեւ թարթիչավոր (կեղծ բազմաշերտ) 1 - թարթիչավոր բջիջ; 2 - շողացող թարթիչ; 3 - ներդիր (փոխարինող) բջիջ;
Integumentary epithelium (սխեմա ըստ Ալեքսանդրովսկայայի). B - խորանարդ; B - գլանաձեւ (սյունակ); G - բազմաշերտ գլանաձեւ թարթիչավոր (կեղծ բազմաշերտ) 1 - թարթիչավոր բջիջ; 2 - շողացող թարթիչ; 3 - ներդիր (փոխարինող) բջիջ;
 Շիճուկային թաղանթների միաշերտ թիթեղային էպիթելը (պլեվրա և պերիտոնե) կոչվում է մեզոթելիա, արյունատար անոթների ներքին պատերը, թոքերի ալվեոլները և աչքերի ցանցաթաղանթը կոչվում են էնդոթելի։
Շիճուկային թաղանթների միաշերտ թիթեղային էպիթելը (պլեվրա և պերիտոնե) կոչվում է մեզոթելիա, արյունատար անոթների ներքին պատերը, թոքերի ալվեոլները և աչքերի ցանցաթաղանթը կոչվում են էնդոթելի։
 Միաշերտ տափակ էպիթելի (մեզոթելիում) օմենտի շիճուկային թաղանթից Նշումներ. 1 - բջջային սահմաններ; 2 - միջուկներ mesotheliocytes; 3 - երկմիջուկային բջիջներ; 4 - «լյուկներ» Դեղը բարակ թաղանթ է, որի հիմքում ընկած է չամրացված շարակցական հյուսվածքը, երկու կողմից ծածկված միաշերտ թիթեղային էպիթելով՝ մեսոթելիումով։ Մեզոթելային բջիջները հարթ են, մեծ, թեթև ցիտոպլազմով և կլորացված միջուկներով։ Բջիջների եզրագծերն ունեն ատամնավոր տեսք և ակնհայտորեն հակադրվում են սև արծաթի նստվածքով: Որոշ տեղերում խցերի միջև կան փոքր անցքեր՝ ԼՈՒԿԵՐ։
Միաշերտ տափակ էպիթելի (մեզոթելիում) օմենտի շիճուկային թաղանթից Նշումներ. 1 - բջջային սահմաններ; 2 - միջուկներ mesotheliocytes; 3 - երկմիջուկային բջիջներ; 4 - «լյուկներ» Դեղը բարակ թաղանթ է, որի հիմքում ընկած է չամրացված շարակցական հյուսվածքը, երկու կողմից ծածկված միաշերտ թիթեղային էպիթելով՝ մեսոթելիումով։ Մեզոթելային բջիջները հարթ են, մեծ, թեթև ցիտոպլազմով և կլորացված միջուկներով։ Բջիջների եզրագծերն ունեն ատամնավոր տեսք և ակնհայտորեն հակադրվում են սև արծաթի նստվածքով: Որոշ տեղերում խցերի միջև կան փոքր անցքեր՝ ԼՈՒԿԵՐ։
 Միաշերտ խորանարդ էպիթելի հայտնաբերվում է գեղձերի խողովակներում, երիկամների խողովակներում, վահանաձև գեղձի ֆոլիկուլներում: Միաշերտ պրիզմատիկ էպիթելը հայտնաբերվում է աղիքների, ստամոքսի, արգանդի, ձվաբջիջների լորձաթաղանթում, ինչպես նաև լյարդի, ենթաստամոքսային գեղձի արտազատվող խողովակներում։ Պրիզմատիկ էպիթելի տեսակները ներառում են լիմբիկ (աղիքային էպիթել) և գեղձային (ստամոքսի էպիթել):
Միաշերտ խորանարդ էպիթելի հայտնաբերվում է գեղձերի խողովակներում, երիկամների խողովակներում, վահանաձև գեղձի ֆոլիկուլներում: Միաշերտ պրիզմատիկ էպիթելը հայտնաբերվում է աղիքների, ստամոքսի, արգանդի, ձվաբջիջների լորձաթաղանթում, ինչպես նաև լյարդի, ենթաստամոքսային գեղձի արտազատվող խողովակներում։ Պրիզմատիկ էպիթելի տեսակները ներառում են լիմբիկ (աղիքային էպիթել) և գեղձային (ստամոքսի էպիթել):
 Բազմաշար թարթիչավոր էպիթելը բջիջների ազատ ծայրերում կրում է 20270 տատանվող թարթիչներ։ Նրանց շարժումների օգնությամբ շնչառական ուղիներից և կանանց սեռական օրգաններից հեռացվում են պինդ կամ հեղուկ օտար մասնիկները։
Բազմաշար թարթիչավոր էպիթելը բջիջների ազատ ծայրերում կրում է 20270 տատանվող թարթիչներ։ Նրանց շարժումների օգնությամբ շնչառական ուղիներից և կանանց սեռական օրգաններից հեռացվում են պինդ կամ հեղուկ օտար մասնիկները։
 Պարզ էպիթել A - շերտավոր B - միաշերտ խորանարդ C - գլանաձև D - գլանաձև թարթիչավոր D - զգայական հատուկ զգայական ելքերով E - գեղձային էպիթել, որը պարունակում է գավաթային բջիջներ, որոնք արտազատում են լորձ
Պարզ էպիթել A - շերտավոր B - միաշերտ խորանարդ C - գլանաձև D - գլանաձև թարթիչավոր D - զգայական հատուկ զգայական ելքերով E - գեղձային էպիթել, որը պարունակում է գավաթային բջիջներ, որոնք արտազատում են լորձ
 Շերտավորված էպիթելը բաղկացած է մի քանի բջիջների շերտերից.
Շերտավորված էպիթելը բաղկացած է մի քանի բջիջների շերտերից.
 Ներկված էպիթելիա (սխեմա ըստ Ալեքսանդրովսկայայի)՝ բազմաշերտ՝ D - հարթ (փեղկավոր) չկերատինացնող՝ բազալային շերտի 1 բջիջ; փշոտ շերտի 2 բջիջ; 3 - մակերեսային շերտի բջիջ; E - հարթ (squamous) keratinizing: 1 - basal շերտ; 2 - փշոտ; 3 - հատիկավոր; 4 փայլուն; 5 եղջյուրավոր; G - անցումային `բազային շերտի 1 բջիջ; 2 - միջանկյալ շերտի բջիջներ; 3 - բջիջները integumentary շերտի. Պինդ սլաքը ցույց է տալիս չամրացված շարակցական հյուսվածքը, գծավոր սլաքը ցույց է տալիս գավաթի բջիջը
Ներկված էպիթելիա (սխեմա ըստ Ալեքսանդրովսկայայի)՝ բազմաշերտ՝ D - հարթ (փեղկավոր) չկերատինացնող՝ բազալային շերտի 1 բջիջ; փշոտ շերտի 2 բջիջ; 3 - մակերեսային շերտի բջիջ; E - հարթ (squamous) keratinizing: 1 - basal շերտ; 2 - փշոտ; 3 - հատիկավոր; 4 փայլուն; 5 եղջյուրավոր; G - անցումային `բազային շերտի 1 բջիջ; 2 - միջանկյալ շերտի բջիջներ; 3 - բջիջները integumentary շերտի. Պինդ սլաքը ցույց է տալիս չամրացված շարակցական հյուսվածքը, գծավոր սլաքը ցույց է տալիս գավաթի բջիջը
 Չկերատինացված էպիթելը հայտնաբերվում է աչքերի, կերակրափողի և հեշտոցի եղջերաթաղանթում: Կերատինացնող էպիթելը կազմում է մաշկի մակերեսային շերտը՝ էպիդերմիսը, այն նաև գծում է բերանի խոռոչի, կոկորդի, կերակրափողի լորձաթաղանթը։ Այս տեսակի էպիթելիումը բաղկացած է աստիճանաբար կերատինացող բջիջների չորս շերտերից՝ ամենախոր շերտը աճող շերտն է, այն բաղկացած է կենդանի բջիջներից, որոնք չեն կորցրել միտոզելու ունակությունը։ հատիկավոր շերտ շերտ corneum շերտ corneum կազմված եղջյուրավոր թեփուկներից
Չկերատինացված էպիթելը հայտնաբերվում է աչքերի, կերակրափողի և հեշտոցի եղջերաթաղանթում: Կերատինացնող էպիթելը կազմում է մաշկի մակերեսային շերտը՝ էպիդերմիսը, այն նաև գծում է բերանի խոռոչի, կոկորդի, կերակրափողի լորձաթաղանթը։ Այս տեսակի էպիթելիումը բաղկացած է աստիճանաբար կերատինացող բջիջների չորս շերտերից՝ ամենախոր շերտը աճող շերտն է, այն բաղկացած է կենդանի բջիջներից, որոնք չեն կորցրել միտոզելու ունակությունը։ հատիկավոր շերտ շերտ corneum շերտ corneum կազմված եղջյուրավոր թեփուկներից


 Շերտավորված տափակ ոչ կերատինացված էպիթելը և գեղձային էպիթելը շան կերակրափողի մի հատվածից լորձաթաղանթով պատված է շերտավորված տափակ ոչ կերատինացված էպիթելով, որը գտնվում է ալիքաձև նկուղային թաղանթի վրա: Նշումներ `1 - նկուղային թաղանթ; 2 - բազալ շերտ; 3 - փշոտ շերտ; 4 - մակերեսային շերտ; 5 - չամրացված կապ հյուսվածք; 6 - լորձաթաղանթների արտազատիչ հատվածներ; 7 - գեղձերի արտազատիչ խողովակներ Լորձաթաղանթի չամրացված շարակցական հյուսվածքում կան բարդ ճյուղավորված գլանային ալվեոլային լորձաթաղանթներ։ Արտազատող խողովակները նման են տարբեր հարթություններում կտրված խողովակների:
Շերտավորված տափակ ոչ կերատինացված էպիթելը և գեղձային էպիթելը շան կերակրափողի մի հատվածից լորձաթաղանթով պատված է շերտավորված տափակ ոչ կերատինացված էպիթելով, որը գտնվում է ալիքաձև նկուղային թաղանթի վրա: Նշումներ `1 - նկուղային թաղանթ; 2 - բազալ շերտ; 3 - փշոտ շերտ; 4 - մակերեսային շերտ; 5 - չամրացված կապ հյուսվածք; 6 - լորձաթաղանթների արտազատիչ հատվածներ; 7 - գեղձերի արտազատիչ խողովակներ Լորձաթաղանթի չամրացված շարակցական հյուսվածքում կան բարդ ճյուղավորված գլանային ալվեոլային լորձաթաղանթներ։ Արտազատող խողովակները նման են տարբեր հարթություններում կտրված խողովակների:
 Շերտավորված անցումային էպիթելը գծում է միզուղիների լորձաթաղանթները: Քանի որ նրանց խոռոչների ծավալը փոխվում է այս օրգանների աշխատանքի ընթացքում, էպիթելի շերտի հաստությունը ենթարկվում է ձգման և սեղմման։
Շերտավորված անցումային էպիթելը գծում է միզուղիների լորձաթաղանթները: Քանի որ նրանց խոռոչների ծավալը փոխվում է այս օրգանների աշխատանքի ընթացքում, էպիթելի շերտի հաստությունը ենթարկվում է ձգման և սեղմման։
 Շան միզապարկ. Անցումային էպիթելի Նշումներ՝ I - լորձաթաղանթ՝ 1 - անցումային էպիթելիա; 2 - սեփական ափսե; 3 - ենթամեկուսային հիմք; II - մկանային թաղանթ `4 - ներքին երկայնական շերտ; 5 - միջին շրջանաձեւ շերտ; 6 - արտաքին երկայնական շերտ; 7 - չամրացված միացնող հյուսվածքի շերտեր; 8 - անոթներ; III - արտաքին պատյան
Շան միզապարկ. Անցումային էպիթելի Նշումներ՝ I - լորձաթաղանթ՝ 1 - անցումային էպիթելիա; 2 - սեփական ափսե; 3 - ենթամեկուսային հիմք; II - մկանային թաղանթ `4 - ներքին երկայնական շերտ; 5 - միջին շրջանաձեւ շերտ; 6 - արտաքին երկայնական շերտ; 7 - չամրացված միացնող հյուսվածքի շերտեր; 8 - անոթներ; III - արտաքին պատյան
 Գեղձային էպիթելի էպիթելային հյուսվածքի բջիջները ունակ են սինթեզելու այլ օրգանների գործառույթների իրականացման համար անհրաժեշտ ակտիվ նյութեր (սեկրեցիա, հորմոն): Գաղտնիքներ արտադրող էպիթելը կոչվում է գեղձային, իսկ նրա բջիջները՝ սեկրետորային բջիջներ (գրանուլոցիտներ):
Գեղձային էպիթելի էպիթելային հյուսվածքի բջիջները ունակ են սինթեզելու այլ օրգանների գործառույթների իրականացման համար անհրաժեշտ ակտիվ նյութեր (սեկրեցիա, հորմոն): Գաղտնիքներ արտադրող էպիթելը կոչվում է գեղձային, իսկ նրա բջիջները՝ սեկրետորային բջիջներ (գրանուլոցիտներ):
 Խցուկներ Էնդոկրին էնդո - ներսում, կրիո - առանձին Նրանք զրկված են արտազատվող խողովակներից, նրանց ակտիվ նյութերը (հորմոնները) արյան մեջ են մտնում մազանոթների միջոցով (վահանաձև գեղձ, հիպոֆիզ, մակերիկամներ): Էկզոկրին էքսոն արտաքինից Գաղտնիքներն արտազատվում են գեղձերի կողմից, որոնք ունեն ծորաններ (կաթնագեղձեր, քրտինք, թքագեղձեր):
Խցուկներ Էնդոկրին էնդո - ներսում, կրիո - առանձին Նրանք զրկված են արտազատվող խողովակներից, նրանց ակտիվ նյութերը (հորմոնները) արյան մեջ են մտնում մազանոթների միջոցով (վահանաձև գեղձ, հիպոֆիզ, մակերիկամներ): Էկզոկրին էքսոն արտաքինից Գաղտնիքներն արտազատվում են գեղձերի կողմից, որոնք ունեն ծորաններ (կաթնագեղձեր, քրտինք, թքագեղձեր):

 Խցուկների տեսակները (ըստ արտազատման մեթոդի) հոլոկրին խցուկներ (որոնցում անընդհատ տեղի է ունենում բջիջների ամբողջական քայքայում և սեկրեցիա)։ Օրինակ, մաշկի ճարպային գեղձը; ապոկրին գեղձեր (բջջի մի մասը քայքայված է)՝ մակրոապոկրին (քանդված է գեղձի գագաթնակետը) միկրոապոկրին (միկրովիլի գագաթային մասերն առանձնացված են)։ Ապոկրինային գեղձերը կաթնագեղձերն ու քրտինքի գեղձերն են: մերոկրին (որում գեղձի բջիջները չեն ոչնչացվում): Այս տեսակի գեղձերը ներառում են՝ թքագեղձեր, ենթաստամոքսային գեղձեր, ստամոքսի գեղձեր, էնդոկրին գեղձեր։
Խցուկների տեսակները (ըստ արտազատման մեթոդի) հոլոկրին խցուկներ (որոնցում անընդհատ տեղի է ունենում բջիջների ամբողջական քայքայում և սեկրեցիա)։ Օրինակ, մաշկի ճարպային գեղձը; ապոկրին գեղձեր (բջջի մի մասը քայքայված է)՝ մակրոապոկրին (քանդված է գեղձի գագաթնակետը) միկրոապոկրին (միկրովիլի գագաթային մասերն առանձնացված են)։ Ապոկրինային գեղձերը կաթնագեղձերն ու քրտինքի գեղձերն են: մերոկրին (որում գեղձի բջիջները չեն ոչնչացվում): Այս տեսակի գեղձերը ներառում են՝ թքագեղձեր, ենթաստամոքսային գեղձեր, ստամոքսի գեղձեր, էնդոկրին գեղձեր։
 Աջակցող-տրոֆիկ (միացնող հյուսվածքներ) Ø արյուն Ø ավիշ Ø աճառ հյուսվածք Ø ոսկրային հյուսվածք Այս տեսակը ներառում է հյուսվածքներ, որոնք կազմում են օրգանների կմախքը և կենդանու ամբողջ մարմինը, դրանք կազմում են մարմնի ներքին միջավայրը:
Աջակցող-տրոֆիկ (միացնող հյուսվածքներ) Ø արյուն Ø ավիշ Ø աճառ հյուսվածք Ø ոսկրային հյուսվածք Այս տեսակը ներառում է հյուսվածքներ, որոնք կազմում են օրգանների կմախքը և կենդանու ամբողջ մարմինը, դրանք կազմում են մարմնի ներքին միջավայրը:
 Հյուսվածքների ընդհանուր մորֆոլոգիական առանձնահատկությունը ոչ միայն բջիջների, այլև միջբջջային նյութի բաղադրության մեջ առկայությունն է։ Հիմնական գործառույթներն են մարմնի օժանդակ, տրոֆիկ, կենսաբանական պաշտպանությունը։
Հյուսվածքների ընդհանուր մորֆոլոգիական առանձնահատկությունը ոչ միայն բջիջների, այլև միջբջջային նյութի բաղադրության մեջ առկայությունն է։ Հիմնական գործառույթներն են մարմնի օժանդակ, տրոֆիկ, կենսաբանական պաշտպանությունը։

 Մեզենխիմը ամենապրիմիտիվ հյուսվածքն է, որը հանդիպում է միայն սաղմերում։ Այն կառուցված է սինցիցիումի սկզբունքով (սաղմնային ցանցով կապված պրոցեսային բջիջների մի շարք), որոնց ընդմիջումներում առկա է դոնդողանման միջբջջային նյութ։
Մեզենխիմը ամենապրիմիտիվ հյուսվածքն է, որը հանդիպում է միայն սաղմերում։ Այն կառուցված է սինցիցիումի սկզբունքով (սաղմնային ցանցով կապված պրոցեսային բջիջների մի շարք), որոնց ընդմիջումներում առկա է դոնդողանման միջբջջային նյութ։


 Լիմֆը բաղկացած է հեղուկ մասից՝ լիմֆոպլազմից և լիմֆոցիտների ձևավորված տարրերից. - Ծայրամասային ավիշ (ավիշ մազանոթներ և անոթներ դեպի ավշային հանգույցներ) - միջանկյալ ավիշ (անոթային ավիշ՝ ավշային հանգույցներով անցնելուց հետո) - կենտրոնական ավիշ (ավշային և աջ ավշային լիմֆատիկ): խողովակներ)
Լիմֆը բաղկացած է հեղուկ մասից՝ լիմֆոպլազմից և լիմֆոցիտների ձևավորված տարրերից. - Ծայրամասային ավիշ (ավիշ մազանոթներ և անոթներ դեպի ավշային հանգույցներ) - միջանկյալ ավիշ (անոթային ավիշ՝ ավշային հանգույցներով անցնելուց հետո) - կենտրոնական ավիշ (ավշային և աջ ավշային լիմֆատիկ): խողովակներ)
 Աճառ Հիալինային կամ ապակենման աճառ (հոդային մակերեսներին, կողերի ծայրերին, քթի միջնապատում, շնչափողում և բրոնխներում) Էլաստիկ աճառ ( ականջի խոռոչում, էպիգլոտտում, արտաքին լսողական խողովակում) Թելքավոր աճառ (միջողնաշարային սկավառակներ, հանգույցներ) ջլերից մինչև ոսկորներ)
Աճառ Հիալինային կամ ապակենման աճառ (հոդային մակերեսներին, կողերի ծայրերին, քթի միջնապատում, շնչափողում և բրոնխներում) Էլաստիկ աճառ ( ականջի խոռոչում, էպիգլոտտում, արտաքին լսողական խողովակում) Թելքավոր աճառ (միջողնաշարային սկավառակներ, հանգույցներ) ջլերից մինչև ոսկորներ)
 Hyaline աճառ 1 - perichondrium; 2 աճառային գոտի երիտասարդ աճառային բջիջներով; 3 - հիմնական նյութ; 4 - բարձր տարբերակված աճառային բջիջներ; 5 - աճառային բջիջների իզոգեն խմբեր; 6 պարկուճ աճառ բջիջներ; 7 Բազոֆիլ աղացած նյութ աճառի բջիջների շուրջ
Hyaline աճառ 1 - perichondrium; 2 աճառային գոտի երիտասարդ աճառային բջիջներով; 3 - հիմնական նյութ; 4 - բարձր տարբերակված աճառային բջիջներ; 5 - աճառային բջիջների իզոգեն խմբեր; 6 պարկուճ աճառ բջիջներ; 7 Բազոֆիլ աղացած նյութ աճառի բջիջների շուրջ
 ականջի առաձգական աճառ՝ 1 պերիխոնդրիում; 2 - երիտասարդ աճառ բջիջներ; 3 - աճառային բջիջների իզոգեն խմբեր; 4 - առաձգական մանրաթելեր
ականջի առաձգական աճառ՝ 1 պերիխոնդրիում; 2 - երիտասարդ աճառ բջիջներ; 3 - աճառային բջիջների իզոգեն խմբեր; 4 - առաձգական մանրաթելեր
 Թելքավոր աճառ ջիլը տիբիային կցվելու վայրում. 1 - ջիլ բջիջներ; 2 - աճառ բջիջները
Թելքավոր աճառ ջիլը տիբիային կցվելու վայրում. 1 - ջիլ բջիջներ; 2 - աճառ բջիջները
 Ոսկրային հյուսվածքը (textus osseus) շարակցական հյուսվածքի հանքայնացված տեսակ է, որը չոր զանգվածում պարունակում է գրեթե 70% անօրգանական միացություններ, հիմնականում կալցիումի ֆոսֆատ: Կատարում է օժանդակ, մեխանիկական, կալցիումի աղերի պահեստ և ներքին օրգանների պաշտպանիչ գործառույթներ։
Ոսկրային հյուսվածքը (textus osseus) շարակցական հյուսվածքի հանքայնացված տեսակ է, որը չոր զանգվածում պարունակում է գրեթե 70% անօրգանական միացություններ, հիմնականում կալցիումի ֆոսֆատ: Կատարում է օժանդակ, մեխանիկական, կալցիումի աղերի պահեստ և ներքին օրգանների պաշտպանիչ գործառույթներ։
 Կախված կառուցվածքային առանձնահատկություններից՝ առանձնանում են ոսկրային հյուսվածքի երկու տեսակ՝ կոպիտ մանրաթելային շերտավոր Կոպիտ մանրաթելային սաղմնային ոսկրային հյուսվածք է՝ մեծ քանակությամբ բջջային տարրերով և կապոցներով հավաքված կոլագենի մանրաթելերի անկանոն դասավորությամբ։ Այնուհետև, կոպիտ մանրաթելային հյուսվածքը փոխարինվում է շերտավոր ոսկրային հյուսվածքով, որը բաղկացած է բջիջներից և ոսկրային թիթեղներից, որոնք ունեն որոշակի տարածական կողմնորոշում, և դրանցում առկա բջիջներն ու կոլագենի մանրաթելերը պարփակված են հանքայնացված ամորֆ նյութի մեջ: Կմախքի հարթ և խողովակային ոսկորներից կազմված կոմպակտ և սպունգանման նյութ է ձևավորվում շերտավոր ոսկրային հյուսվածքից։
Կախված կառուցվածքային առանձնահատկություններից՝ առանձնանում են ոսկրային հյուսվածքի երկու տեսակ՝ կոպիտ մանրաթելային շերտավոր Կոպիտ մանրաթելային սաղմնային ոսկրային հյուսվածք է՝ մեծ քանակությամբ բջջային տարրերով և կապոցներով հավաքված կոլագենի մանրաթելերի անկանոն դասավորությամբ։ Այնուհետև, կոպիտ մանրաթելային հյուսվածքը փոխարինվում է շերտավոր ոսկրային հյուսվածքով, որը բաղկացած է բջիջներից և ոսկրային թիթեղներից, որոնք ունեն որոշակի տարածական կողմնորոշում, և դրանցում առկա բջիջներն ու կոլագենի մանրաթելերը պարփակված են հանքայնացված ամորֆ նյութի մեջ: Կմախքի հարթ և խողովակային ոսկորներից կազմված կոմպակտ և սպունգանման նյութ է ձևավորվում շերտավոր ոսկրային հյուսվածքից։
 Խողովակային ոսկորի կառուցվածքի սխեման `1 - պերիոստեում; 2 - հարսյան ալիք; 3 - ներդիր համակարգ; 4 - հարսյան համակարգ; 5 - ոսկրային թիթեղների արտաքին ընդհանուր համակարգ; 6 - արյան անոթներ; 7 Volkman ալիք; 8 - կոմպակտ ոսկոր; 9 - սպունգային ոսկոր; 10 - ոսկրային թիթեղների ներքին ընդհանուր համակարգ
Խողովակային ոսկորի կառուցվածքի սխեման `1 - պերիոստեում; 2 - հարսյան ալիք; 3 - ներդիր համակարգ; 4 - հարսյան համակարգ; 5 - ոսկրային թիթեղների արտաքին ընդհանուր համակարգ; 6 - արյան անոթներ; 7 Volkman ալիք; 8 - կոմպակտ ոսկոր; 9 - սպունգային ոսկոր; 10 - ոսկրային թիթեղների ներքին ընդհանուր համակարգ
 Հատուկ հատկություններով շարակցական հյուսվածք՝ ցանցաթաղանթային ճարպային պիգմենտային լորձաթաղանթ Այն բնութագրվում է որոշակի տեսակի բջիջների գերակշռությամբ.
Հատուկ հատկություններով շարակցական հյուսվածք՝ ցանցաթաղանթային ճարպային պիգմենտային լորձաթաղանթ Այն բնութագրվում է որոշակի տեսակի բջիջների գերակշռությամբ.
 Ցանցային հյուսվածքը ձևավորվում է ցանցային բջիջներով և դրանց ածանցյալներով՝ ցանցաթելերով։ Ցանցային հյուսվածքը ձևավորում է արյունաստեղծ օրգանների ստրոմա և ստեղծում միկրոմիջավայր արյան բջիջների և մակրոֆագների համար: Ճարպային հյուսվածքը ճարպային բջիջների հավաքածու է, որն ապահովում է օրգանիզմում լիպիդների սինթեզն ու կուտակումը։ Տարբերակել սպիտակ և շագանակագույն ճարպային հյուսվածքը: Պիգմենտային շարակցական հյուսվածքը չամրացված թելքավոր շարակցական հյուսվածք է՝ պիգմենտային բջիջների զգալի գերակշռությամբ: Պիգմենտային հյուսվածքի օրինակ է աչքի ծիածանաթաղանթի և քորոիդային հյուսվածքը: Լորձաթաղանթային շարակցական հյուսվածքն առկա է միայն սաղմնային շրջանում, այն հանդիպում է բազմաթիվ օրգաններում, հատկապես մաշկի տակ։ Լորձաթաղանթի հյուսվածքի օրինակ է պտղի պորտալարի հյուսվածքը:
Ցանցային հյուսվածքը ձևավորվում է ցանցային բջիջներով և դրանց ածանցյալներով՝ ցանցաթելերով։ Ցանցային հյուսվածքը ձևավորում է արյունաստեղծ օրգանների ստրոմա և ստեղծում միկրոմիջավայր արյան բջիջների և մակրոֆագների համար: Ճարպային հյուսվածքը ճարպային բջիջների հավաքածու է, որն ապահովում է օրգանիզմում լիպիդների սինթեզն ու կուտակումը։ Տարբերակել սպիտակ և շագանակագույն ճարպային հյուսվածքը: Պիգմենտային շարակցական հյուսվածքը չամրացված թելքավոր շարակցական հյուսվածք է՝ պիգմենտային բջիջների զգալի գերակշռությամբ: Պիգմենտային հյուսվածքի օրինակ է աչքի ծիածանաթաղանթի և քորոիդային հյուսվածքը: Լորձաթաղանթային շարակցական հյուսվածքն առկա է միայն սաղմնային շրջանում, այն հանդիպում է բազմաթիվ օրգաններում, հատկապես մաշկի տակ։ Լորձաթաղանթի հյուսվածքի օրինակ է պտղի պորտալարի հյուսվածքը:
 Մկանային հյուսվածք Մկանային հյուսվածքը ծագումով և կառուցվածքով տարասեռ հյուսվածքների խումբ է, որը միավորված է մեկ և նրա համար հիմնական ֆունկցիոնալ հատկանիշով` կծկվելու ունակությամբ, որն ուղեկցվում է թաղանթային ներուժի փոփոխությամբ: Կախված կծկվող օրգանելների՝ միոֆիբրիլների մորֆոֆունկցիոնալ առանձնահատկություններից, մկանային հյուսվածքները բաժանվում են.
Մկանային հյուսվածք Մկանային հյուսվածքը ծագումով և կառուցվածքով տարասեռ հյուսվածքների խումբ է, որը միավորված է մեկ և նրա համար հիմնական ֆունկցիոնալ հատկանիշով` կծկվելու ունակությամբ, որն ուղեկցվում է թաղանթային ներուժի փոփոխությամբ: Կախված կծկվող օրգանելների՝ միոֆիբրիլների մորֆոֆունկցիոնալ առանձնահատկություններից, մկանային հյուսվածքները բաժանվում են.

 Նյարդային հյուսվածքը մարմնում ապահովում է տարբեր հյուսվածքների և օրգանների փոխազդեցության կարգավորում և հաղորդակցություն շրջակա միջավայրի հետ՝ հիմնված գրգռման և իմպուլսների անցկացման վրա՝ մասնագիտացված կառույցների միջոցով: Նյարդային հյուսվածքը կառուցված է նյարդային բջիջներից (նեյրոցիտներ, նեյրոններ) և նեյրոգլիայից։ Նեյրոնը մասնագիտացված հյուսվածքի հիմնական կառուցվածքային բաղադրիչն է: Կատարում է իմպուլս վարելու գործառույթ: Նեյրոգլիան կատարում է տրոֆիկ, սահմանազատող, աջակցող, արտազատող և պաշտպանիչ գործառույթներ։
Նյարդային հյուսվածքը մարմնում ապահովում է տարբեր հյուսվածքների և օրգանների փոխազդեցության կարգավորում և հաղորդակցություն շրջակա միջավայրի հետ՝ հիմնված գրգռման և իմպուլսների անցկացման վրա՝ մասնագիտացված կառույցների միջոցով: Նյարդային հյուսվածքը կառուցված է նյարդային բջիջներից (նեյրոցիտներ, նեյրոններ) և նեյրոգլիայից։ Նեյրոնը մասնագիտացված հյուսվածքի հիմնական կառուցվածքային բաղադրիչն է: Կատարում է իմպուլս վարելու գործառույթ: Նեյրոգլիան կատարում է տրոֆիկ, սահմանազատող, աջակցող, արտազատող և պաշտպանիչ գործառույթներ։
 Նեյրոններում՝ մարմնում կամ պերիկարիոնում, նյարդային մանրաթելեր ձևավորող գործընթացները և նյարդային վերջավորությունները մեկուսացված են։ Նեյրոններն ունեն մասնագիտացված պլազմալեմա, որն ունակ է գրգռում իրականացնել պրոցեսներից դեպի մարմին և դրանից դեպի գործընթաց՝ ապաբևեռացման պատճառով: Նյարդային պրոցեսները ֆունկցիոնալորեն բաժանվում են՝ աքսոնը կամ նևրիտը նեյրոնի մարմնից այլ նեյրոն կամ աշխատանքային օրգանի հյուսվածքներ տարածում է դեպի մկաններ, դենդրիտների գեղձերը ընկալում են գրգռվածություն, ձևավորում իմպուլս և այն տանում դեպի նեյրոն։ նեյրոն
Նեյրոններում՝ մարմնում կամ պերիկարիոնում, նյարդային մանրաթելեր ձևավորող գործընթացները և նյարդային վերջավորությունները մեկուսացված են։ Նեյրոններն ունեն մասնագիտացված պլազմալեմա, որն ունակ է գրգռում իրականացնել պրոցեսներից դեպի մարմին և դրանից դեպի գործընթաց՝ ապաբևեռացման պատճառով: Նյարդային պրոցեսները ֆունկցիոնալորեն բաժանվում են՝ աքսոնը կամ նևրիտը նեյրոնի մարմնից այլ նեյրոն կամ աշխատանքային օրգանի հյուսվածքներ տարածում է դեպի մկաններ, դենդրիտների գեղձերը ընկալում են գրգռվածություն, ձևավորում իմպուլս և այն տանում դեպի նեյրոն։ նեյրոն
 Նյարդային բջիջի կառուցվածքը `1 - մարմին (պերիկարիոն); 2 միջուկ; 3 - դենդրիտներ; 4 - նեյրիտներ; 5, 8 - միելինային պատյան; 7 գրավ; 9 հանգույցի ընդհատում; 10 - լեմմոցիտ; 11 - նյարդային վերջավորություններ
Նյարդային բջիջի կառուցվածքը `1 - մարմին (պերիկարիոն); 2 միջուկ; 3 - դենդրիտներ; 4 - նեյրիտներ; 5, 8 - միելինային պատյան; 7 գրավ; 9 հանգույցի ընդհատում; 10 - լեմմոցիտ; 11 - նյարդային վերջավորություններ
 ՌՆԹ-ի բոլոր տեսակները. ՌՆԹ-ի կառուցվածքը և գործառույթները. ՌՆԹ բջիջների տեսակներն ու տեսակները
ՌՆԹ-ի բոլոր տեսակները. ՌՆԹ-ի կառուցվածքը և գործառույթները. ՌՆԹ բջիջների տեսակներն ու տեսակները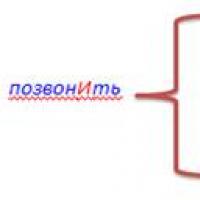 Ո՞ր բառում է ճիշտ ընդգծված ընդգծված ձայնավորը նշանակող տառը:
Ո՞ր բառում է ճիշտ ընդգծված ընդգծված ձայնավորը նշանակող տառը: Ռազմաբժշկական ակադեմիա
Ռազմաբժշկական ակադեմիա Ներկայացում «զրույց» թեմայով կետադրական և ուղղագրական տաքացում
Ներկայացում «զրույց» թեմայով կետադրական և ուղղագրական տաքացում Անպարկեշտ խաղաղություն. ինչպես ազդեց Բրեստ-Լիտովսկի պայմանագիրը Ռուսաստանի պատմության ընթացքի վրա
Անպարկեշտ խաղաղություն. ինչպես ազդեց Բրեստ-Լիտովսկի պայմանագիրը Ռուսաստանի պատմության ընթացքի վրա Պիտեր I-ի քաղաքական բարեփոխումները
Պիտեր I-ի քաղաքական բարեփոխումները Հայրենական մեծ պատերազմի տարիներին ԽՍՀՄ զինվորական շքանշանների և մեդալների լուսանկարներ
Հայրենական մեծ պատերազմի տարիներին ԽՍՀՄ զինվորական շքանշանների և մեդալների լուսանկարներ